Автономний (вегетативний) відділ периферійної нервової системи
Загальна характеристика автономного (вегетативного) відділу нервової системи
Ще з часів К. Біша (1800) загальновизнано, що вегетативні функції, або функції рослинного життя (дихання, травлення, розмноження, виділення і тощо) регулюються структурами вегетативної нервової системи. Дж. Ленглі в 1889 році поділив усю вегетативну нервову систему на два основні відділи – парасимпатичний та симпатичний. Окремо ним була виділена так звана ентеральна система, або кишкове сплетення.
Сучасна Міжнародна номенклатура пропонує не використовувати термін автономна (вегетативна) нервова система, щоб уникнути багатозначності. Натомість пропонується термін автономний відділ; автономна частина периферійної нервової системи, divisio autonomica (pars autonomica systematis nervosi peripherici), в якій розрізняють симпатичну частину (pars sympathica) та парасимпатичну частину (pars parasympathica). Слід, однак, зазначити, що у вітчизняній науковій та навчальній літературі термін “вегетативна нервова система” є поширеним і тому більш зрозумілим. Крім того, в останні роки завдяки дослідженням А. Д. Ноздрачова (1983,1989) у межах вегетативної нервової системи запропоновано виділяти ще одну частину (систему, відділ) – метасимпатичну.
За сучасним визначенням автономна (вегетативна) нервова система – це частина нервової системи, що виконує функції регуляції постійності внутрішнього середовища організму (гомеостазу) і пристосування до умов навколишнього середовища, яке постійно змінюється (гомеокінезу). Регуляція гомеостазу включає підтримку на потрібному для організму рівні біохімічних, фізико-хімічних, ферментативних та інших констант, порушення яких проявляється не тільки численними вегетативними, але й соматичними дисфункціями. Регуляція гомеокінезу включає забезпечення різних форм діяльності (розумової, емоційної, фізичної) та реалізацію біологічних мотивацій цілісного організму. Порушення адекватних гомеокінетичних реакцій змінює поведінку людини, призводить до дезадаптації і зрештою до виникнення захворювання або загибелі організму.
На даний час симпатична частина автономної (вегетативної) нервової системи розглядається як система тривоги, мобілізації захисних сил і ресурсів для активної взаємодії з факторами середовища, тобто як ерготропна система. Парасимпатична частина, як і метасимпатична, призначена для відновлення і підтримки гомеостазу, порушеного внаслідок активності організму, тобто здійснює трофотропний вплив. При цьому метасимпатична частина автономної (вегетативної) нервової системи реалізує цю функцію регіонально, тобто в окремому органі чи його частині.
Поняття “автономна” нервова система – відносне поняття, оскільки діяльність цієї частини нервової системи хоч і спрямована на процеси регуляції роботи внутрішніх органів і найчастіше не підпорядковується свідомому і вольовому контролю, але функціонування вегетативної нервової системи відбувається згідно з роботою сенсорних, рухових та інтелектуальних систем мозку.
Вегетативна нервова система – це комплексне поняття. В кожному відділі спинного і головного мозку, а також на периферії є скупчення нейронів, які належать до вегетативної нервової системи. За аналогією з руховими системами мозку правильніше було б говорити про вегетативні системи певного відділу мозку і периферії. У даний час найбільш визнаним є уявлення, згідно з яким вегетативна нервова система – це центральні сегментні структури, розташовані у стовбурі головного мозку і у спинному мозку, а також периферійні структури, тобто зв’язані з ними через передвузлові волокна, вегетативні вузли або ганглії, а також завузлові (постгангліонарні) волокна, що відходять від вузлів до органа.
До центральних сегментних структур вегетативної нервової системи належать парасимпатичні ядра ІІІ, VІI, IX, X пар черепних нервів, парасимпатичні нейрони крижового відділу спинного мозку, а також симпатичні нейрони, що містяться в бокових рогах тораколюмбального відділу спинного мозку. Всі ці центральні сегментні осередки (центри) внаслідок особливості їх організації та закономірності функціонування є справжніми автономними центрами.
Крім цього, є вегетативні центри регуляції вісцеральних функцій, які контролюють діяльність усіх трьох частин вегетативної (автономної) системи. Ці центри розміщені в довгастому мозку і в мості (центри регуляції окремих систем організму, наприклад, дихальний центр, судиноруховий центр), у ретикулярній формації стовбура (моноамінергічна система), в гіпоталамусі та інших структурах лімбічної системи (інтегральні центри регуляції діяльності внутрішніх органів, судин і різних гомеостатичних процесів), а також у мозочку, базальних ядрах і структурах нової кори півкуль великого мозку. Всі ці утворення запропоновано називати надсегментними структурами вегетативної нервової системи, або вищими вегетативними центрами, основним завданням яких є організація діяльності функціональних систем, що відповідають за регуляцію психічних, соматичних і вісцеральних функцій.
Свого часу І. П. Павлов вказував, що “чим досконаліша нервова система живого організму, тим вона більш централізована, тим вищий відділ її все більшою й більшою мірою є розпорядником і розподілювачем усієї діяльності організму, незважаючи на те, що це яскраво і відкрито не виступає. Адже нам може здаватись, що багато функцій у вищих тварин іде зовсім поза впливом великих півкуль, а насправді це не так. Цей вищий відділ контролює всі явища, які відбуваються в тілі”.
Отже, узагальнюючи вищесказане, вегетативна нервова система є не ізольованою автономною системою, а спеціальним відділом єдиної нервової системи, що підкоряється її вищим відділам, включаючи і кору півкуль великого мозку. Саме тому, як і в соматичній нервовій системі, у вегетативній можна розрізняти центральний і периферійний відділи.
До центрального відділу належать описані вище осередки (симпатичні і парасимпатичні сегментні центри) у спинному мозку і в стовбурі головного мозку, а також надсегментні центри (спільні для всіх частин вегетативної нервової системи) в головному мозку, а до периферійного – нервові вузли, нерви, сплетення і периферійні нервові закінчення.
Найсуттєвішими морфологічними відмінностями автономної (вегетативної) нервової системи від соматичної нервової системи є:
1) розміщення осередків її парасимпатичної та симпатичної частин у певних місцях ЦНС (сегментні центри);
2) розташування аксона другого нейрона і тіла третього нейрона простої рефлекторної дуги поза межами ЦНС. Тіла третіх нейронів скупчуються у вигляді вегетативних вузлів. Тому волокна автономної (вегетативної) нервової системи, що несуть імпульс від спинного або головного мозку до вузла, називають передвузловими (прегангліонарними), а волокна, що несуть імпульс від вузла до органа, – завузловими (постгангліонарними).
За ступенем віддалення від ЦНС і за топографією вегетативні вузли поділяють на:
1) розміщені з боків хребтового стовпа вузли першої черги, або прихребтові вузли (вузли симпатичного стовбура);
2) розміщені спереду хребтового стовпа вузли другої черги, або передхребтові вузли;
3) розміщені поблизу органів або в тканинах внутрішніх органів вузли ІІІ черги (відповідно, позата внутрішньоорганні).
Вузли І та ІІ черги є симпатичними; вузли ІІІ черги переважно є парасимпатичними. Основна частина передвузлових вегетативних волокон – це мієлінові тонкі волокна діаметром не більше 5 мкм, а завузлові волокна ще тонші і не мають мієлінової оболонки. Еферентні волокна соматичної нервової системи в основному є товстими мієліновими волокнами великого (12–14 мкм) діаметра. Всі вегетативні волокна мають низьку збудливість, низьку лабільність, а також невелику швидкість проведення нервового імпульсу, яка не перевищує 18 і 3 м/с відповідно для переді завузлових волокон.
У закінченнях всіх передвузлових симпатичних і парасимпатичних нейронів, а також у закінченнях більшості завузлових парасимпатичних волокон виділяється нейротрансмітер ацетилхолін. Крім того, частина завузлових симпатичних волокон, що іннервують потові залози, також здійснюють передачу за допомогою ацетилхоліну.
Нейротрансмітером у завузлових симпатичних закінченнях (за винятком нервів потових залоз і симпатичних вазодилататорів) є норадреналін.
Слід зазначити, що адренергічні нейрони мають дуже довгі тонкі аксони, які розгалужуються в органах і утворюють густі сплетення. Загальна довжина таких аксонних терміналів може досягати 30 см. По ходу терміналів є численні розширення – варикози, в яких синтезується, накопичується і виділяється нейротрансмітер. З приходом імпульсу норадреналін одночасно виділяється з багатьох розширень, діючи відразу на велику площу тканини.
Більшість органів мають подвійну іннервацію симпатичними та парасимпатичними завузловими волокнами. Винятком є утвори симпатоадреналової або хромафінної системи (мозкова речовина надниркових залоз та параганглії), що мають тільки симпатичну іннервацію, яка здійснюється лише передвузловими волокнами. Це пов’язано з тим, що мозкова речовина надниркових залоз і параганглії за ембріологічним походженням є зміненими симпатичними вузлами і розвиваються з симпатобластів, які мігрують із закладок вузлів симпатичного стовбура.
Для нейронів метасимпатичної частини автономного відділу периферійної нервової системи характерний широкий набір нейротрансмітерів, тому серед них розрізняють холінергічні, адренергічні, серотонінергічні, пептидергічні та інші нейрони.
Серотонін, взаємодіючи із специфічними серотоніновими рецепторами, здатний діяти безпосередньо на міоцити судин, матки, бронхів і шлунково-кишкового тракту.
При взаємодії з пуринорецепторами тонкої і товстої кишок АТФ викликає їх розслаблення.
У механізмі кишкової перистальтики пуринергічні нейрони є головною антагоністичною гальмуючою системою щодо збуджуючої холінергічної системи. Пуринергічні нейрони беруть участь у здійсненні низхідного гальмування, в механізмі рецептивної релаксії шлунка, розслаблення стравохідного та анального сфінктерів.
Гістамін, що продукується гістамінергічними метасимпатичними нейронами, викликає розслаблення міоцитів судин, збільшення проникності капілярів, збільшення скоротливості міоцитів бронхів і матки, посилює моторну та секреторну функції шлунка.
Важливим трансмітером метасимпатичних нейронів є ГАМК, яка здійснює гальмівний вплив на міжнейронну передачу збудження в інтрамуральних вегетативних вузлах.
Субстанція Р (пептид) є нейротрансмітером чутливих метасимпатичних нейронів в інтрамуральних вузлах. Особливо багато цих нейронів знаходиться у травному тракті.
Найбільш суттєва відмінність між симпатичною та парасимпатичною частинами вегетативної нервової системи полягає:
1) у різній довжині передвузлових та завузлових волокон (симпатична частина має короткі передвузлові та довгі завузлові волокна, парасимпатична – навпаки, тобто парасимпатичні вузли розташовані набагато ближче до робочого органа);
2) у характері дії нейротрансмітерів, що виділяються на кінцях завузлових волокон (норадреналін одночасно з місцевою дією зумовлює і загальну, ацетилхолін – тільки місцеву і, крім того, швидко руйнується холінестеразою);
3) у тому, що аксон нейронів сегментних центрів парасимпатичної частини контактує (ефект мультиплікації) з меншим числом нейронів вегетативного вузла, ніж аксон сегментних центрів симпатичної. Усе це зумовлює більш дифузний і генералізований симпатичний та локальний парасимпатичний ефект.
В органах з подвійною вегетативною іннервацією спостерігається взаємодія функцій симпатичних і парасимпатичних нервів у формі антагонізму спричинюваних ефектів, якщо ці нерви подразнюються окремо. Наприклад, подразнення симпатичних нервів викликає розширення зіниць, гальмування перистальтики, розслаблення сфінктерів, розширення бронхів і вінцевих артерій, посилення і прискорення серцевих скорочень. Подразнення парасимпатичних нервів призводить до звуження зіниць, посилення перистальтики, закриття сфінктерів, звуження бронхів і вінцевих артерій, до сповільнення та ослаблення серцебиття.
Слід підкреслити, що в умовах здійснення вегетативних і соматичних функцій організм як єдина цілісна система широко використовує як симпатичні, так і парасимпатичні нервові структури, тому їх функціональний антагонізм і синергізм є двома складовими одного й того ж процесу регуляції гомеостазу та гомеокінезу.
Вказані вище анатомічні особливості вегетативної нервової системи дозволяють здійснювати переключення вісцеральних аферентних сигналів на еферентні клітини на різних рівнях, у тому числі на рівні прихребтових, передхребтових або інтрамуральних вузлів, на спінальному, бульбарному і кірковому рівнях. При цьому на всіх перелічених рівнях існує спеціальний інтернейронний апарат, що здійснює первинну обробку сенсорних сигналів, а це збільшує можливість використання рефлексів як основи регуляції діяльності внутрішніх органів. Потрібно зазначити, що діяльність вегетативної нервової системи тісно пов’язана з діяльністю рухових систем мозку. Тому нерідко вісцеральна аферентація викликає і вегетативні, і рухові рефлекси.
Нині виділяють такі види рефлексів вегетативної нервової системи, як вісцеро-вісцеральні, вісцеро-соматичні, вісцеро-сенсорні, вісцеро-дермальні, дермовісцеральні і сомато-вісцеральні.
Вісцеро-вісцеральний рефлекс полягає в тому, що рефлексогенною зоною є один будь-який внутрішній орган, а роль ефектора виконує інший орган, діяльність якого може бути загальмована або, навпаки, прискорена. Класичним прикладом є рефлекс Гольца, при якому механічне подразнення брижі тонкої кишки викликає сповільнення частоти серцевих скорочень. Іншим прикладом служить подразнення рецепторів травного тракту, що супроводжується послабленням тонусу м’яза-звужувача зіниці. Подразнення рефлексогенної зони в ділянці sinus caroticus (у місці роздвоєння загальної сонної артерії) викликає зміну інтенсивності дихання, рівня кров’яного тиску, частоти серцевих скорочень.
Аксон-рефлекс є різновидом вісцеро-вісцерального рефлексу. Він реалізується без участі тіла нейрона – збудження виникає в одній гілці аксона, потім переходить на іншу і по ній спрямовується до виконавчого органа, викликаючи відповідну реакцію. Цей механізм, імовірно, лежить в основі появи судинної реакції (розширення судини) при подразненні шкірних больових рецепторів, наприклад, при реалізації рефлексу, що отримав назву “місцевий дермографізм”.
Вісцеро-соматичний рефлекс виникає у тому випадку, коли рефлексогенною зоною є який-небудь один внутрішній орган, а в ролі ефектора виступає один або кілька скелетних м’язів, які можуть відповідати скороченням або розслабленням. Прикладом такої реакції може служити гальмування загальної рухової активності організму при подразненні чутливих закінчень синокаротидної зони, а також скорочення м’язів черевної стінки або посмикування кінцівок при подразненні рецепторів травного тракту.
Вісцеро-сенсорний рефлекс виникає в тому випадку, коли рефлексогенною зоною є який-небудь внутрішній орган, а роль ефектора (виконавця) виконують одна або декілька сенсорних систем. Найчастіше йдеться про підвищення дотикової або больової чутливості. При цьому зона підвищеного сприйняття обмежується ділянкою шкіри, що іннервується сегментом спинного мозку, до якого надходять імпульси від вісцерального органа, який подразнюють. Механізм цього явища пов’язаний з тим, що вісцеральні і шкірні чутливі волокна конвергують на одних і тих самих нейронах спинно-таламічного шляху. При подальшій передачі сенсорних сигналів до кори півкуль великого мозку у проміжних структурах втрачається специфічність інформації, внаслідок чого кора зв’язує виникаючі збудження з подразненням певної ділянки шкірної поверхні. Проявом вісцеро-сенсорних рефлексів є відомі клінічні спостереження, згідно з якими при захворюванні внутрішніх органів виникає підвищення дотикової і больової чутливості певних (залежно від виду органа) ділянок шкіри. Ці болі названі відбитими (віддзеркаленими), а ділянки їх прояву – зонами Захар’їна – Геда.
Вісцеро-сенсорні рефлекси подібні до вісцеровісцеральних і вісцеро-соматичних рефлексів. Але вони, як правило, виникають при тривалій і сильній дії подразника на даний внутрішній орган. Це означає, що при збільшенні сили подразника виникає ймовірність того, що паралельно з вісцеро-вісцеральними рефлексами будуть виникати вісцеро-соматичні і вісцеро-сенсорні рефлекси.
Вісцеро-дермальний рефлекс – варіант двох видів рефлексів: вісцеро-вісцерального і вісцеро-сенсорного. Рефлекс полягає у тому, що подразнення внутрішніх органів одночасно супроводжується зміною потовиділення (і як наслідок цього – зміною електричного опору або електропровідності шкіри) і зміною шкірної чутливості. Цей рефлекс лежить в основі шкірно-гальванічного рефлексу.
Рефлексогенною зоною сомато-вісцерального і дермо-вісцерального рефлексів є скелетні м’язи або ділянки шкіри, а ефектором – міоцити судин та інших органів, а також залозиста тканина або серцевий м’яз. Ці рефлекси проявляються тим, що при подразненні деяких ділянок поверхні тіла виникають судинні реакції і змінюється функція певних внутрішніх органів. Це явище лежить в основі рефлексотерапії – одного із перспективних напрямків клінічної та профілактичної медицини.
Парасимпатична частина автономного відділу периферійної нервової системи
Парасимпатична частина автономного відділу периферійної нервової системи (або, за традиційною термінологією, – периферійний відділ парасимпатичної частини вегетативної нервової системи) представлений передвузловими волокнами, що йдуть ускладі ІІІ, VII, ІХ і Х пар черепних нервів і тазових нутрощевих нервів, парасимпатичними вузлами (позаорганними та внутрішньоорганними), а також відповідними завузловими волокнами.
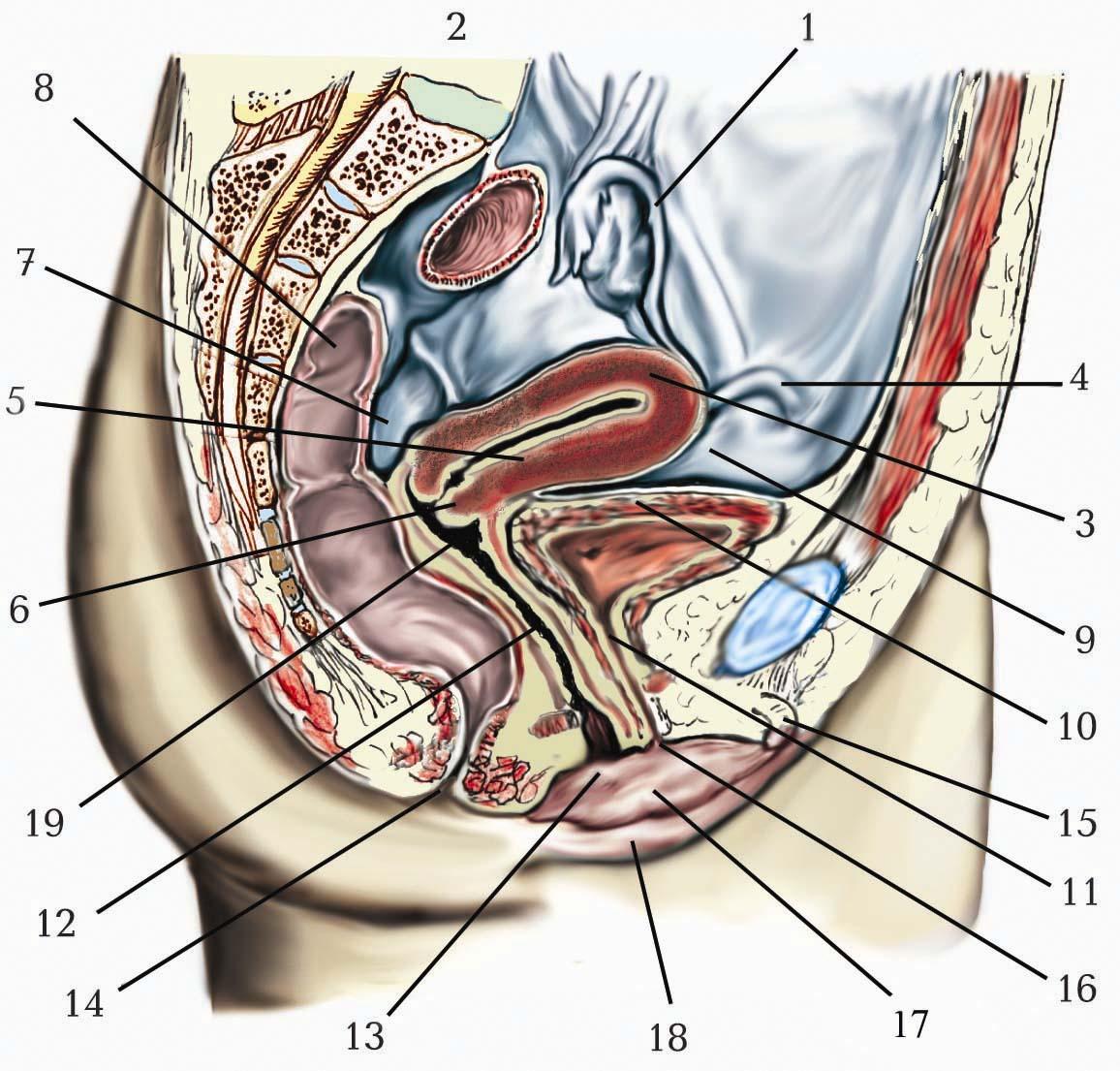
Позаорганні парасимпатичні вузли розміщені поблизу органів, що іннервуються. До них належать п’ять великих парасимпатичних вузлів голови (війковий, крило-піднебінний, піднижньощелепний, під’язиковий, вушний), а також парасимпатичні тазові вузли.
До цих вузлів підходять три типи нервів чи корінців: чутливі, симпатичні та парасимпатичні. Перших два типи волокон проходять транзитом, при цьому чутливі волокна несуть інформацію до мозку, а симпатичні (завузлові) – до органів, що ними іннервуються. Волокна парасимпатичних корінців закінчуються на нейронах вузлів. Аксони цих нейронів утворюють завузлові волокна, які досягають органа, що іннервується. Периферійні гілки описаних вузлів утворені трьома різновидами волокон (чутливими, симпатичними і парасимпатичними).
Війковий вузол (ganglion ciliare) має непостійну форму та величину (його довжина в середньому складає 2 мм). Цей вузол лежить у товщі жирової тканини у задньому відділі очної ямки на зовнішній поверхні зорового нерва (відступаючи на 7–8 мм від заднього полюса очного яблука). Війковий вузол має три корінці:
1) парасимпатичний корінець (radix parasympathica) – утворений гілкою окорухового нерва до війкового вузла, r. n. oculomotorius ad ganglion ciliare;
2) симпатичний корінець (radix sympathica);
3) чутливий корінець (radix sensoria) – утворений сполучною гілкою носовійкового нерва з війковим вузлом (r. communicans n. nasociliaris cum ganglio ciliare).
Завузлові парасимпатичні волокна цього вузла у складі коротких війкових нервів (nn. ciliares breves) прямують до очного яблука, пронизують склеру й іннервують війковий м’яз та м’яз-звужувач зіниці.
Крило-піднебінний вузол(ganglion pterygopalatinum) розміром близько 5 мм, розміщений у крилопіднебінній ямці збоку від клино-піднебінного отвору. Крило-піднебінний вузол має три корінці:
1) парасимпатичний корінець (radix parasympathica) – утворений великим кам’янистим нервом (n. petrosus major);
2) симпатичний корінець (radix sympathica) – утворений глибоким кам’янистим нервом (n. petrosus profundus);
3) чутливий корінець (radix sensoria) – утворений вузловими гілками верхньощелепного нерва (rr. ganglionares n. maxillaris). Завузлові парасимпатичні волокна цього вузла (у складі гілок верхньощелепного нерва) забезпечують секреторну іннервацію залоз слизової оболонки порожнини носа, рота, глотки та сльозової залози.
Піднижньощелепний вузол (ganglion submandibulare) розміром близько 3 мм, лежить на піднижньощелепній слинній залозі під n. lingualis. Піднижньощелепний вузол має три корінці:
1) парасимпатичний корінець (radix parasympathica) – утворений барабанною струною (chorda tympani);
2) симпатичний корінець (radix sympathica);
3) чутливий корінець (radix sensoria) – утворений вузловими гілками піднижньощелепного нерва (rr. ganglionares n. mandibularis). Завузлові парасимпатичні волокна цього вузла забезпечують секреторну іннервацію піднижньощелепної та під’язикової слинних залоз.
Під’язиковий вузол (ganglion sublinguale) дуже мінливий за будовою, інколи розміщується на бічній поверхні під’язикової слинної залози, але частіше є скупченням нейронів по ходу гілок n. lingualis до під’язикової залози. Під’язиковий вузол має три корінці:
1) парасимпатичний корінець (radix parasympathica) – утворений барабанною струною (chorda tympani);
2) симпатичний корінець (radix sympathica);
3) чутливий корінець (radix sensoria) – утворений вузловими гілками піднижньощелепного нерва (rr. ganglionares n. mandibularis). Завузлові парасимпатичні волокна цього вузла забезпечують секреторну іннервацію під’язикової залози.
Вушний вузол (ganglion oticum) розміром близько 3 мм, розміщений у підскроневій ямці присередньо від n.mandibularis під овальним отвором. Вушний вузол має три корінці:
1) парасимпатичний корінець (radix parasympathica) – утворений малим кам’янистим нервом (n. petrosus minor);
2) симпатичний корінець (radix sympathica);
3) чутливий корінець (radix sensoria) – утворений вузловими гілками піднижньощелепного нерва (rr. ganglionares n. mandibularis). Завузлові парасимпатичні волокна цього вузла забезпечують секреторну іннервацію привушної слинної залози.
Тазові вузли (ganglia pelvica) розміщені у складі нижнього підчеревного сплетення та інших тазових вегетативних сплетень. Кожен з них має три корінці:
1) симпатичний корінець (radix sympathica);
2) парасимпатичний корінець (radix parasympathica) – утворений тазовими нутрощевими нервами (nn. splanchnici pelvici);
3) чутливий корінець (radix sensoria) – утворений nn. splanchnici pelvici. Nn. splanchnici pelvici йдуть від передніх гілок ІІ–ІV крижових нервів (більша частина їхніх волокон є передвузловими парасимпатичними, менша – чутливими). Nn. splanchnici pelvici забезпечують парасимпатичну та чутливу іннервацію органів малого таза та кишкового тракту нижче низхідної ободової кишки.
Крім описаних великих парасимпатичних вузлів, які мають відповідні корінці, є дрібні парасимпатичні вузли автономних сплетень (gg. plexuum autonomicum). Ці парасимпатичні вузли розміщені у позаорганних та внутрішньоорганних вегетативних сплетеннях паренхіматозних органів шиї, грудної, черевної і тазової порожнин. Передвузлові парасимпатичні волокна йдуть до них у складі гілок блукаючого нерва і тазових нутрощевих нервів.
Крім того, раніше виділяли дрібні інтрамуральні парасимпатичні вузли, що входять до складу інтрамуральних нервових сплетень, які розміщені в різних шарах стінок трубчастих органів. Нині їх розглядають як складову метасимпатичної частини автономного відділу периферійної нервової системи.
Симпатична частина автономного відділу периферійної нервової системи
Симпатична частина автономного відділу периферійної нервової системи (або, за традиційною термінологією, – периферійний відділ симпатичної частини вегетативної нервової системи) представлений передвузловими та завузловими волокнами, а також прихребтовими (вузлами симпатичного стовбура) та передхребтовими (нутрощевими вузлами, або вузлами нутрощевих сплетень) симпатичними вузлами.
Як було відзначено вище, симпатична частина автономного відділу ПНС призначена для здійснення ерготропного впливу на діяльність внутрішніх органів, судин і на процеси, що беруть участь у ресинтезі АТФ, а також для проявлення адаптаційно-трофічного впливу на організм людини.
У симпатичних вузлах відбувається мультиплікація: одне передвузлове волокно передає сигнал до багатьох завузлових нейронів, відбувається “розмноження” сигналу. Завдяки цьому вплив симпатичної системи набуває генералізованого характеру. Далі в симпатичних вузлах може відбуватися змикання місцевих (периферійних) рефлексів – здійснюється майже автономна регуляція діяльності органа.
Прихребтові симпатичні вузли розміщені з боків хребтового стовпа у вигляді парного ланцюга – симпатичного стовбура (truncus sympathicus). Симпатичний стовбур загалом складається з вузлів симпатичного стовбура (ganglia trunci sympathici), з’єднаних між собою короткими міжвузловими гілками (rr. interganglionares). Кількість вузлів симпатичного стовбура, відповідно до поділу хребтового стовпа, наближається до кількості хребців даної ділянки (за винятком шийного і куприкового відділів, де вона значно менша). Куприковий відділ представлений загальним для обох симпатичних стовбурів маленьким непарним вузлом (ganglion impar), в якому вони закінчуються.
Вузли симпатичного стовбура зв’язані з сусідніми спинномозковими нервами за допомогою сполучних гілок (rr. communicantes). Розрізняють білі і сірі сполучні гілки. Білі сполучні гілки (rr. communicantes albi) складаються з мієлінових передвузлових волокон, утворених аксонами нейронів nucleus intermediolateralis бічних рогів тораколюмбального відділу спинного мозку. Ці волокна йдуть у складі передніх корінців відповідних спинномозкових нервів (у межах від І грудного до ІІІ поперекового) і поблизу їх з’єднання із задніми корінцями відокремлюються, прямуючи до найближчого вузла симпатичного стовбура. Від цих вузлів симпатичного стовбура до всіх спинномозкових нервів відходять сірі сполучні гілки (rr. communicantes grisei), які складаються з безмієлінових завузлових волокон, утворених аксонами нейронів цих вузлів. Увійшовши в кожний спинномозковий нерв, завузлові симпатичні волокна поширюються з його гілками на периферію, досягаючи робочих органів.
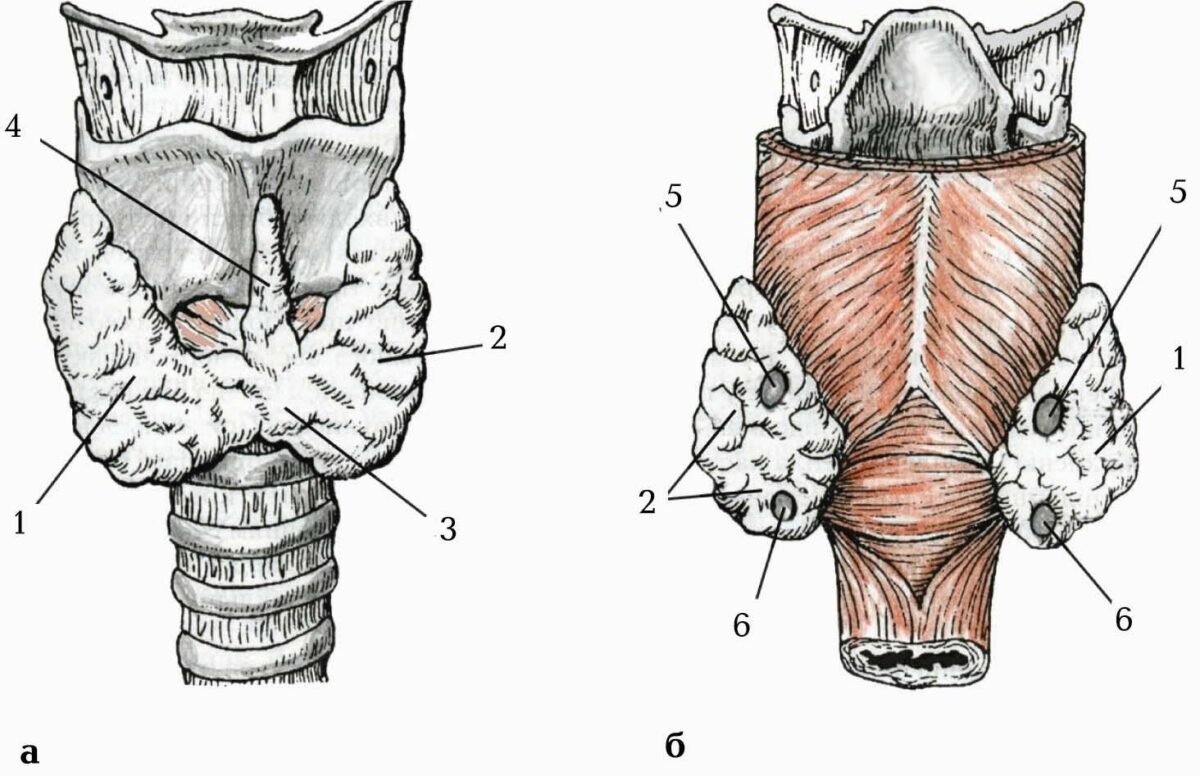
Шийний відділ симпатичного стовбура вкритий fascia prevertebralis і містить три вузли: верхній, середній і нижній.
Верхній шийний вузол (ganglion cervicale superius) найбільший з усіх вузлів симпатичного стовбура (завдовжки до 2 см і більше), має веретеноподібну форму і розміщується спереду поперечних відростків ІІ–ІІІ шийних хребців позаду a. carotis interna. Від верхнього шийного вузла відходять: яремний нерв, внутрішній сонний нерв, зовнішні сонні нерви, гортанно-глоткові гілки, верхній шийний серцевий нерв.
Внутрішній сонний нерв (n. caroticus internus) разом з шийною частиною a. carotis interna піднімається до canalis caroticus і, супроводжуючи артерію в каналі, утворює навколо неї внутрішнє сонне сплетення, яке потім переходить у печеристе сплетення (оточує печеристу частину a. carotis interna). З останнього починаються сплетення, які оточують гілки мозкової частини a. carotis interna.
Зовнішні сонні нерви (nn. carotici externi) мають вигляд 2–3 тонких стовбурців, які утворюють порівняно незначне зовнішнє сонне сплетення. Останнє дає початок сплетенням, які супроводять гілки зовнішньої сонної артерії.
Гортанно-глоткові гілки (rr. laryngopharyngei) частково йдуть з n. laryngeus superior (гілка n. vagus) до гортані; частково йдуть разом з глотковими гілками n. vagus et n. glossopharyngeus і беруть участь в утворенні глоткового сплетення у стінці глотки; частково йдуть разом зі стравохідними гілками n. vagus, приймаючи участь в утворенні стравохідного сплетення.
Верхній шийний серцевий нерв (n. cardiacus cervicalis superior) спускається присередньо від симпатичного стовбура, перетинає ззаду a. thyroidea inferior і прямує до серцевого сплетення: з правого боку – уздовж truncus brachiocephalicus, з лівого – уздовж a. carotis communis sinistra.
Середній шийний вузол (ganglion cervicale medium) має незначні розміри і лежить на рівні VI шийного хребця. Від цього вузла відходить середній шийний серцевий нерв (n. cardiacus cervicalis medius), який, спускаючись позаду a. carotis communis, іде до серцевого сплетення.
Нижній шийний вузол (ganglion cervicale inferior) розміщується позаду a. subclavia, біля початку a. vertebralis. Цей вузол часто зливається з верхнім грудним вузлом у шийно-грудний або зірчастий вузол (ganglion cervicothoracicum seu ganglion stellatum). Нижній шийний вузол віддає нижній шийний серцевий нерв (n. cardiacus cervicalis inferior), який спускається до серцевого сплетення, та хребтовий нерв (n. vertebralis), який утворює навколо a. vertebralis хребтове сплетення.
Грудний відділ симпатичного стовбура вкритий fascia endothoracica i pleura costalis. Він містить 11–12 вузлів, які лежать спереду головок ребер (тільки два нижні – на бічній поверхні тіл хребців).
Грудні вузли (ganglia thoracica) віддають: грудні серцеві гілки, грудні легеневі гілки, стравохідні гілки, великий нутрощевий нерв, малий нутрощевий нерв, найнижчий нутрощевий нерв.
Грудні серцеві гілки (rr. cardiaci thoracici) утворені завузловими симпатичними та аферентними (больовими) волокнами ІІ–IV грудних вузлів; беруть участь в утворенні серцевого сплетення.
Грудні легеневі гілки (rr. pulmonares thoracici) утворені волокнами ІІ–IV грудних вузлів; беруть участь в утворенні легеневого сплетення.
Стравохідні гілки (rr. oesophageales) утворені волокнами ІІ–V грудних вузлів; беруть участь в утворенні стравохідного сплетення.
Великий нутрощевий нерв (n. splanchnicus major) починається від V–IX грудних вузлів корінцями, які, йдучи медіально і донизу по бічній поверхні хребців, з’єднуються один з одним у стовбур. Останній спускається попереду хребтового стовпа і разом з v. azygos проходить через поперекову частину діафрагми у черевну порожнину і входить до складу черевного сплетення. У товщі великого нутрощевого нерва на рівні IX грудного хребця розміщений грудний нутрощевий вузол (ganglion thoracicum splanchnicum).
Малий нутрощевий нерв (n. splanchnicus minor) починається корінцями від X–XI грудних вузлів, іде латеральніше попереднього через діафрагму і досягає черевного сплетення. Значна частина волокон n. splanchnicus minor формує ниркову гілку (r. renalis), яка безпосередньо прямує до ниркового сплетення. До ниркового сплетення від XII грудного вузла йде також найнижчий нутрощевий нерв (n. splanchnicus imus).
Поперековий відділ симпатичного стовбура складається з 4–5 поперекових вузлів (ganglia lumbalia), менших, ніж грудні, розміщених на передній поверхні тіл хребців, уздовж присереднього краю m. psoas major . З правого боку поперекові вузли прикриті нижньою порожнистою веною, з лівого – тягнуться по краю аорти. Від поперекових вузлів у медіальному напрямі відходять поперекові нутрощеві нерви (nn. splanchnici lumbales), які лягають на передню поверхню поперекових хребців, утворюючи сплетення, і прямують далі до нутрощевих сплетень черевної порожнини.
Крижово-куприковий відділ симпатичного стовбура складається з чотирьох пар крижових вузлів (ganglia sacrales) і одного куприкового непарного вузла (ganglion impar). Крижові вузли розміщені на тазовій поверхні крижової кістки медіально від передніх крижових отворів . Чим нижче, тим вузли дрібніші і тим ближче вони лежать до серединної площини. Від останньої пари вузлів відходять каудально міжвузлові гілки до непарного вузла. Непарний вузол має незначні розміри і лежить на передній поверхні тіла І куприкового хребця. Таким чином, обидва ланцюжки симпатичного стовбура внизу змикаються. Від крижових вузлів відходять тонкі крижові нутрощеві нерви (nn. splanchnici sacrales), які беруть участь у формуванні нутрощевих сплетень тазової порожнини.
Метасимпатична частина автономного відділу периферійної нервової системи
За Ноздрачовим А. Д. (1983, 1991, 1997), який увів поняття “метасимпатична система”, дана частина автономного відділу периферійної нервової системи вирізняється відносною простотою. Тут немає ядерних структур і система представлена лише комплексом інтрамуральних дрібновузлових структур, що залягають у стінках порожнистих органів, які мають моторну активність. Ідеться про наявність дрібних вузлів (подібно до внутрішньоорганних вузлів парасимпатичної частини) в серці, бронхах, шлунку, кишці, сечовому міхурі, сечівнику, матці, сім’явиносних шляхах. Відповідно до ділянок іннервації, Ноздрачов пропонує в метасимпатичній частині автономного відділу периферійної нервової системи виділяти кардіальну, респіраторну, ентеральну та інші частини.
На думку Ноздрачова (1991, 1997), метасимпатична частина автономного відділу периферійної нервової системи може, по-перше, здійснювати передачу центральних впливів, оскільки парасимпатичні і симпатичні волокна можуть контактувати з метасимпатичною частиною і тим самим контролювати її вплив на об’єкти управління. По-друге, метасимпатична частина може виконувати роль самостійного інтегруючого утворення, оскільки в ній є готові рефлекторні дуги, що містять аферентні, вставні та еферентні нейрони.
Найбільш вивчена метасимпатична іннервація шлунково-кишкового тракту, в якому описане кишкове сплетення (plexus entericus), що складається з:
– підсерозного сплетення (plexus subserosus);
– м’язово-кишкового сплетення (plexus myentericus) – сплетення Ауербаха;
– підслизового сплетення (plexus submucosus) – сплетення Мейснера.
В кожному із цих сплетень є численні дрібні вузли, в яких виділяють три типи нейронів (за Догелем).
Нейрони І типу є еферентними нейронами, аксон яких безпосередньо контактує з м’язовою клітиною, яку іннервує. Нейрони ІІ типу є аферентними нейронами. Частина аксонів цих нейронів переключається на нейрони І типу. У таких випадках рефлекторна дуга замикається на рівні дрібного вузла (мікроганглія). Друга частина аксонів нейронів ІІ типу йде, згідно з даними Ноздрачова, до прихребтового або передхребтового вузлів, переключаючись у них на інші нейрони. Нарешті, ще одна частина аксонів нейронів ІІ типу, за уявленнями Ноздрачова, досягає нейронів спинного мозку, переключаючись у них на інші нейрони. Отже, аферентна імпульсація, що йде від мікрогангліїв, може замикатися на різних рівнях. Нейрони ІІІ типу, за Догелем, є асоціативними нейронами, за участю яких відбувається формування місцевих рефлекторних дуг.
У шлунково-кишковому тракті метасимпатична частина автономного відділу периферійної нервової системи здійснює регуляцію складних рухів різних відділів кишки, в тому числі ритмічних, маятникоподібних, перистальтичних та антиперистальтичних. Очевидно, що для кожного із перелічених видів рухів існує свій метасимпатичний модуль, активація якого відбувається залежно від складу хімусу в даному відділі кишки. Інакше кажучи, завдяки місцевим (метасимпатичним) рефлекторним дугам, активація яких проходить при збудженні відповідних хемо-, механо-, бароі терморецепторів кишки, можлива регуляція його моторики, що пов’язана з процесом гідролізу і всмоктування поживних речовин у шлунково-кишковому тракті.
Детальне вивчення мікроструктури і функціональної організації мікрогангліїв травного тракту дозволило Ноздрачову сформулювати уявлення про механізми роботи метасимпатичного модуля. Вважають, що кожний модуль містить специфічні для нього нейрони-осцилятори, сенсорні нейрони, мотонейрони та інтернейрони. Ключовим нейроном модуля є нейрон-осцилятор. Він має властивість автоматії, тобто здатність до спонтанного збудження (з певною частотою). При кожному збудженні нейрона-осцилятора його потенціали дії передаються через систему вставних нейронів до мотонейрона, тобто до рухового нейрона, аксон якого контактує з кишковим міоцитом. У кишці, як показано в експериментах Ноздрачова, ефекторні мотонейрони за своєю природою є пуринергічними, тобто виділяють у синаптичних закінченнях АТФ, яка є інгібітором скоротливої активності міоцитів кишечнику. Тому при збудженні ефекторного мотонейрона під дією нейрона-осцилятора скорочення гладких міоцитів стінки кишки (в тому числі і спонтанні) сповільнюються. Відповідно, чим активніший нейрон-осцилятор метасимпатичного модуля, тим виразніше гальмування, яке здійснює мотонейрон на міоцити кишки. Потрібно зазначити, що у свою чергу система “нейрон – осцилятор – мотонейрон” може модулюватися. В організмі існують як мінімум два способи такої модуляції:
а) аферентні нейрони модуля, збуджуючись внаслідок активації їхніх дендритів (рецепторів), можуть змінювати активність мотонейрона, діючи на нього безпосередньо (активація соми через холінергічний синапс) або на його закінчення (аксо-аксональне гальмування), знімаючи гальмівний вплив на міоцити;
б) парасимпатичні і симпатичні завузлові волокна, діючи на вставні нейрони модуля, можуть змінювати збудливість мотонейрона метасимпатичного модуля – підвищувати її або, навпаки, знижувати. Наприклад, щодо кишкового тракту встановлено, що при активації парасимпатичних волокон гальмується активність мотонейрона метасимпатичного модуля. Це знімає гальмування метасимпатичного модуля на міоцити кишки, внаслідок чого здійснюється активація міоцитів. Симпатичні волокна, навпаки, посилюють гальмівну дію пуринергічного мотонейрона на міоцити шлунка чи кишки.
За даними Ноздрачова, метасимпатичні модулі здійснюють місцеву регуляцію не тільки скорочення гладеньких м’язів, але й інтенсивності всмоктування, епітеліальної секреції, продукції гормонів, а також інтенсивності місцевих імунних процесів.
Багаторічні дослідження дозволяють відкинути уявлення про те, що метасимпатична нервова система є лише дифузним парасимпатичним вузлом або третім нейроном в еферентній ланці симпатичного шляху. Очевидно, що метасимпатична система, як складова частина вегетативної нервової системи, є незалежною самостійною системою, функцію якої можна порівняти з мікропроцесором, що розташований поблизу ефекторів. Те, що апарат, необхідний для постійного контролю за кожною з вісцеральних функцій, розміщувати в ЦНС недоцільно, підтверджується таким прикладом: тільки в кишці, за розрахунками Ноздрачова, метасимпатична система має таку ж кількість клітин, що й увесь спинний мозок, а кількість метасимпатичних нейронів на 1 см2 поверхні кишки складає близько 20 000. Існування спеціальних місцевих метасимпатичних механізмів регуляції функцій має певний фізіологічний зміст. Вони збільшують надійність регуляції функцій, ця регуляція може відбуватися навіть у випадку втрати зв’язку з центральними структурами. При цьому ЦНС звільнюється від надлишкової інформації.