Бічні шлуночки. Базальні ядра. Локалізація функцій в корі півкуль великого мозку
Морфологічні основи динамічної локалізації функцій в корі півкуль великого мозку
Питання про локалізацію функцій в корі півкуль великого мозку постало давно і має велике теоретичне значення, оскільки дає уявлення про нервову регуляцію всіх процесів організму та пристосувань його до умов навколишнього середовища, які постійно змінюються. Воно має і вагоме практичне значення для встановлення місць уражень у півкулях великого мозку.
До 1861 року кора великого мозку вважалася функціонально однорідною та полівалентною. Будучи (за висловом І. Гіртля) “седалищем души”, кора в функціональному розумінні повинна була бути неподільною. У 1861 році в Парижі П’єр Поль Брока виступив на засіданні Антропологічного товариства і продемонстрував локалізацію функцій в корі головного мозку людини. Він встановив, що порушення мови (моторна афазія) у одного з його хворих виникла внаслідок ушкодження задньої третини лівої нижньої лобової звивини (цю ділянку кори пізніше назвали “центром Брока”). Це відкриття Брока надало імпульс для вивчення локалізації різноманітних функцій в корі мозку.
В 1864 році Хьюлінгс Джексон вивчав хворих з фокальною епілепсією і пояснив причину виникнення припадків подразненням кори передцентральної звивини. Фріч і Гітциг у 1870 році були першими дослідниками, які показали, що електрична стимуляція кори лобової частки півкуль великого мозку у собак призводить до контрлатеральних рухів кінцівок. Але всі перелічені дослідження не мали морфологічного підґрунтя. Засновником вчення про морфологічні основи локалізації функцій в корі великого мозку став видатний український анатом, завідувач кафедри нормальної анатомії Київського університету Св. Володимира, професор Володимир Олексійович Бец. Професор В. О. Бец у 1874 році вперше відкрив і описав у корі gyrus precentralis i lobulus paracentralis гігантські пірамідні клітини (які з того часу у всьому світі називають “клітинами Беца”). У своїй роботі “О подробностях строения мозговой корки человека” (1880) В. О. Бец писав: “В июне 1874 г. мною было опубликовано исследование корки мозга животных, обезьян и преимущественно человека, по которому в местах, соответствующих двигательным центрам Фрича и Гитцига, были найдены особые большие нервные клетки, никем до сих пор не замеченные, имеющие характер клеток спинного мозга и которые я назвал гигантскими клетками… Клетки эти и волокна имеют все несомненные свойства так называемых двигательных клеток и, несомненно, соединяются с нервными волокнами”.
Історія неврології свідчить, що прогрес у спробі визначити і локалізувати кіркові функції був повільним і сповненим протиріч. Із часом в корі півкуль були описані численні цитоархітектонічні поля, але вирішення питання про їх співвідношення з конкретними функціями виявилося утрудненим. Тому вчення про локалізацію функцій у корі великого мозку весь час розвивалося у взаємодії двох протилежних концепцій – антилокалізаціонізму, або еквіпотенціалізму, який відкидає локалізованість функцій у корі, і вузького локалізаційного психоморфологізму, який намагався локалізувати в обмежених ділянках мозку окремі психічні якості.
Розробка основ сучасного нового погляду на локалізацію функцій в корі півкуль великого мозку пов’язана з ім’ям І. П. Павлова, який створив вчення про аналізатори та вчення про динамічну локалізацію функцій.
Павлов розглядав мозкову кору як суцільну сприймаючу поверхню, яка складається з аналізаторів (точніше, їх центральних кінців – мозкових клітин). Аналізатори (зоровий, слуховий, руховий та ін.) не тільки сприймають і диференціюють подразнення, які надходять у мозкову кору із зовнішнього і внутрішнього середовища, але й поєднують їх одне з одним, тобто здійснюють не тільки аналіз, але й синтез.
За І. П. Павловим, кірковий кінець аналізатора (мозковий центр) містить центральну частину, або ядро, де відбувається точне диференціювання подразнень і здійснюється найвищий аналіз і синтез, і периферичну розсіяну ділянку. Периферійні частини різних сприймаючих ділянок кори (кіркових кінців аналізаторів) перекривають одна одну. При ураженні ядра кіркового кінця аналізатора розсіяні елементи можуть до певної міри компенсувати втрачену функцію ядра. Підсумком цих досліджень Павлова стало вчення про динамічну локалізацію функцій в корі мозку, яке припускає можливість участі одних і тих же нервових структур у забезпеченні різних функцій.
Концептуальний підхід Павлова знайшов свій подальший розвиток у вченні про нервові центри і кіркові поля. В сучасній неврології виділяють три типа кіркових полів: первинні, вторинні і третинні. Первинні поля (ядра кіркових кінців аналізаторів за І. П. Павловим) – це проекційні зони, які відповідають тим цитоархітектонічним полям, де закінчуються чутливі провідні шляхи.
Вторинні поля (периферичні відділи кіркових кінців аналізаторів за І. П. Павловим) розміщені поблизу первинних. У цих зонах, безпосередньо не пов’язаних з провідними шляхами, теж відбувається певна обробка інформації, яка надходить у первинні поля. Третинні поля займають решту, тобто більшу частину кори. Це асоціативні зони, які розміщені в місцях перекриття окремих аналізаторів.
Первинні поля (ядра аналізаторів) звичайно мають відносно невеликі розміри та певні анатомічні межі. Що стосується третинних полів, то вони більш розширені і дифузні. З приводу цього слід відмітити, що несподівані результати були отримані при вивченні прозопагнозії (нездатності впізнавати обличчя). Було з’ясовано, що це порушення супроводжується дуже малою кількістю інших неврологічних симптомів, а ділянка асоціативної кори, яка відповідає за цю інформацію, розміщена на нижній поверхні обох потиличних часток і розповсюджується вперед до внутрішньої поверхні скроневих часток. Може здатися, що непропорційно велика кількість мозкових ресурсів витрачається на дуже обмежені задачі. Однак слід мати на увазі, що можливість впізнавання людей як індивідуумів дуже цінна для такого високосоціального біологічного виду, як людина. Подібні приклади свідчать про спеціалізацію мозку людини та велике значення в цьому асоціативних кіркових полів, які незрівнянно перевищують аналогічні зони у тварин. Первинні сенсорні та моторні кіркові поля складають не більше 20 % від загальної поверхні кори людини; решта поверхні зайнята асоціативними полями.
Безумовним стимулом для дослідження локалізації функцій в корі великого мозку став розвиток нейрохірургії. У. Пенфілд і Т. Расмуссен із Монреальського неврологічного інституту в 1950 році, проводячи електростимуляцію кори півкуль у оперованих під місцевою анестезією хворих, одержали нові дані про рухові та чутливі центри мозкової кори. Ними була складена нині широко відома схема, що ілюструє соматичне представництво в первинних сенсорних і первинних моторних кіркових полях, а саме – так звані “чутливий гомункулус” і “руховий гомункулус”. “Чутливий гомункулус” являє собою проекцію на зацентральну звивину протилежної половини тіла людини, причому всі ділянки тіла спроектовані так, що голова «гомункулуса» розташована знизу звивини, а нога зверху. Подібним чином у передцентральній звивині розміщений “руховий гомункулус”.
За допомогою інвазивних методів дослідження були встановлені практично всі первинні кіркові поля (ядра аналізаторів) та деякі вторинні і третинні. Було з’ясовано, що цитоархітектонічні поля 1, 2, 3 (за Бродманом) складають первинне сомато-сенсорне поле, поле 4 і частково 6 – первинне моторне, що поле 17 одержує зорові імпульси, 41 поле – слухові, 43 – смакові і таке інше.
Нові успіхи у вивченні морфологічних основ локалізації функцій в корі півкуль пов’язані із застосуванням високотехнологічних неінвазивних методів дослідження. За допомогою комплексу таких методів (головним з них був метод позитронної емісійної скануючої томографії) Д. Х’юбел та Т. Візел дослідили центральні механізми зору та склали докладну морфологічну схему орієнтаційних і окодомінантних колонок у зоровій корі поля 17 (ділянка острогової борозни), за що в 1981 році одержали Нобелівську премію. Слід зазначити, що, на відміну від клітин інших органів, здатних використовувати різні види «пального», нейрони використовують лише глюкозу крові. Крім того, на відміну від таких тканин, як, наприклад, м’язові, здатних короткочасно функціонувати при відсутності кисню, головний мозок повністю залежить від окислювального метаболізму. Мозок настільки інтенсивно використовує кисень (50 мл за хвилину), що, складаючи всього 2 % загальної маси тіла, поглинає приблизно 20 % кисню, який потрапляє в організм. Таке велике споживання енергії пояснюється необхідністю підтримувати іонні градієнти по обидва боки нейронної мембрани, від чого залежить проведення імпульсів у мільярдах нейронів мозку. Крім того, це споживання енергії відбувається безперервно: інтенсивність метаболізму в мозку відносно постійна вдень і вночі й іноді навіть дещо підвищується під час фази сну зі сновидіннями.
Важливим кроком вперед у дослідженні енергетичного обміну мозку, який призвів до вражаючого розширення та деталізації карти кори великого мозку, став метод, розроблений Л. Соколовим (L. Sokoloff) в Американському національному інституті охорони психічного здоров’я. Цей метод дозволяє візуально визначити інтенсивність енергетичного обміну в клітинах мозку. Нейрони пристосовують споживання глюкози до задоволення своїх метаболічних потреб на даний момент. Отже, в активному стані вони поглинають її скоріше, ніж у спокої. З глюкозою, що була поглинута, звичайно відбуваються швидкі перетворення; її хімічний аналог, 2-дезоксиглюкоза, поглинається клітинами точно так само, але не піддається метаболізму. Якщо ввести в кров дезоксиглюкозу з радіоактивною міткою, то вона відкладеться в нейронах, і швидкість її накопичення буде показником метаболічної активності клітини. Позитронно-емісійна томографія дозволяє виявити за допомогою зовнішніх датчиків наявність дезоксиглюкози або інших речовин, помічених радіоактивними ізотопами, які випромінюють позитрони. Ця перспективна методика дає можливість картування активних структур мозку in vivo у лабораторних тварин або у людини. Наприклад, діючи світловим стимулом (спалах) на праве або ліве око, можна визначити, які саме ділянки мозку одержують аферентацію від того чи іншого ока.
Як установлено, більшість первинних і частина вторинних кіркових полів (кіркових кінців аналізаторів за І. П. Павловим) має певні співвідношення з борознами та звивинами плаща, у зв’язку з чим може бути визначена при дослідженні його рельєфу.
У корі передцентральної звивини та прицентральної часточки розташований кірковий кінець рухового аналізатора (тут сприймається пропріоцептивна чутливість і звідси регулюється діяльність скелетних м’язів). Проекція протилежної половини тіла людини (“руховий гомункулус”) подана догори ногою – у вищих ділянках звивини проектуються рецепторні поля нижньої кінцівки та нижня частина тулуба, а у нижніх відділах – рецепторні поля голови, верхньої кінцівки та верхніх частин тулуба.
У корі зацентральної звивини локалізується кірковий кінець шкірного аналізатора або аналізатора загальної чутливості (больової, температурної, тактильної). У зацентральній звивині, подібно до передцентральної звивини, проекція протилежної половини тіла людини (“чутливий гомункулус”) подана догори ногою.
Специфічним різновидом складної чутливості є стереогноз (тривимірно-просторове почуття), при якому предмет упізнається порівняно з минулим враженням про нього (основна роль при цьому належить суглобово-м’язовій і тактильній чутливості); він пов’язаний з корою верхньої тім’яної часточки домінантної півкулі. У цій ділянці кори розташоване відповідне третинне асоціативне поле.
У ділянці нижньої тім’яної часточки в надкрайовій звивині домінантної півкулі міститься кірковий кінець рухового аналізатора (асоціативне поле), функціональне значення якого полягає у здійсненні всіх цілеспрямованих складних комбінованих рухів, до яких належать також професійні та спортивні рухи.
У задніх відділах середньої лобової звивини розташований кірковий кінець аналізатора співдружнього повороту голови та очей в протилежний бік. Співдружній поворот голови та очей регулюється не тільки пропріоцептивними імпульсами, що йдуть від м’язів очного яблука та м’язів шиї в кору лобової звивини, але й імпульсами, що йдуть від сітківки в поле 17 (sulcus calcarinus потиличної частки).
У людини практично весь мозок задіяний у процесі мовного спілкування (абстрактне мислення пов’язане саме з мовою). Мозок повинен бути непошкодженим, щоб людина без зусиль могла сприймати мову оточуючих і була здатна виразити словами власні думки. Хоч деякі ділянки кори домінантної півкулі є найважливішими для розуміння мови (центр Верніке), а інші – для вимовляння слів (центр Брока), ізольованого мовного центра, мабуть, не існує. Клінічний досвід, однак, показує нерівнозначність окремих ділянок кори в походженні мовних розладів. Втрата моторної або сенсорної функції мови носить назву моторної та сенсорної афазії (від грец. рhasis – мова).
Кірковий кінець рухового аналізатора артикуляції мови (центр Брока) розташований у задніх відділах нижньої лобової звивини домінантної півкулі. При пошкодженні його має місце моторна афазія (втрата рухової програми слова, яка призводить до позбавлення здатності вимовляти склади та слова).
Кірковий кінець слухового аналізатора усної мови (центр Верніке) розташований у задніх відділах верхньої скроневої звивини домінантної півкулі. При пошкодженні його має місце сенсорна афазія (втрата розуміння власної усної мови).
Кірковий кінець рухового аналізатора письмової мови міститься в задніх відділах середньої лобової один приклад асоціативного поля.
У глибині бічної борозни на внутрішній поверхні середньої частини верхньої скроневої звивини в корі поперечних скроневих звивин розміщений кірковий кінець слухового аналізатора.
На присередній поверхні потиличної частки з боків від острогової борозни розміщений кірковий кінець зорового аналізатора.
На нижній поверхні скроневої частки у гачку та частково у підмозолистому полі та морському конику міститься кірковий кінець смакового та нюхового аналізаторів. До первинної смакової кори останнім часом відносять кору лобової та тім’яної покришки, кору острівцевої частки. Описані вище кіркові кінці деяких аналізаторів є в корі півкуль великого мозку не тільки людини, а й тварин. Вони сприймають подразнення із зовнішнього та внутрішнього середовища і складають (за І. П. Павловим) першу сигнальну систему.
Друга сигнальна система є тільки у людини і обумовлена розвитком усної та письмової мови. Мова – центр Верніке.
Кірковий кінець зорового аналізатора письмової мови розташований в кутовій звивині. При пошкодженні його має місце алексія (порушення здатності читати та розуміти написане).
Як уже підкреслювалося раніше, асоціативні кіркові поля лобової, тім’яної, потиличної, скроневої часток півкуль відіграють важливу роль в інтегративній діяльності мозку.
Встановлено, що асоціативні поля, які містяться в корі передніх відділів лобових часток, пов’язані з вищою нервовою діяльністю. Так, ураження передніх двох третин лобових звивин або підлягаючої білої речовини викликає втрату рухової ініціативи і активного мислення.
За клінічними даними інформація з первинних чутливих кіркових полів тім’яної, скроневої, потиличної часток (ядро шкірного, зорового, слухового, нюхового аналізаторів) інтегрується у вторинних і третинних (асоціативних) полях і порівнюється там з раніше накопиченою інформацією або пам’яттю, перетворюючись таким чином у набутий досвід. Переважно цей процес відбувається в домінантній півкулі.
Цитоархітектонічні поля 39, 40 і, можливо 37 поле кутової та надкрайової звивин займають перехідну зону, що з’єднує тактильну та кінестетичну асоціативну ділянку з зоровою і слуховою асоціативними ділянками. У домінантній півкулі ця зона вважається третинним асоціативним полем вищого порядку. Структурні особливості цієї зони аналогічні асоціативним полям лобових часток. У людини кутова та надкрайова звивини займають значно більшу площу в порівнянні з мозком інших приматів і формується досить пізно. Вважають, що тактильна, кінестетична, вестибулярна, зорова, слухова інформація, вже оброблена в первинних і вторинних кіркових полях, інтегрується на вищому рівні в цитоархітектонічних полях 39 і 40. Мабуть, це третинна ділянка (поле) є матеріальним субстратом найскладніших форм людського сприйняття та пізнання.
Базальні (або основні) ядра
Базальні (або основні) ядра (nuclei basales) – це структурно та функціонально неоднорідні утворення сірої речовини у товщі півкуль великого мозку. До базальних ядер належать: смугасте тіло (corpus striatum), огорожа (claustrum) та мигдалеподібне тіло (corpus amygdaloideum). Смугасте тіло складається з хвостатого ядра (nucleus caudatus) і сочевицеподібного ядра (nucleus lentiformis). Сочевицеподібне ядро складається з лушпини (putamen), присередньої блідої кулі (globus pallidus medialis) та бічної блідої кулі (globus pallidus lateralis).
За новою сучасною Міжнародною анатомічною номенклатурою розрізняють новий підрозділ – основна частина кінцевого мозку (pars basalis telencephali), який включає corpus amygdaloideum, claustrum, pallidum ventrale, striatum ventrale, всю передню нюхову частку. Nucleus caudatus, nucleus lentiformis і біла речовина кінцевого мозку об’єднані в основні ядра та структури утворів, nuclei basales et structurae pertinentes. Слід також зазначити, що деякі закордонні підручники стверджують, що substantia nigra i nucleus subthalamicus є частинами базальних ядер кінцевого мозку. Ми в даному підручнику користувались більш традиційним класичним розподілом цих стуктур, беручи до уваги певні розбіжності їх функціональних та морфологічних ознак.
Хвостате ядро та лушпина, хоча і розділені внутрішньою капсулою (capsula interna), є одним ядром і мають однакову гістологічну структуру (щільно розташовані дрібні нейрони та окремі великі мультиполярні нейрони між ними). Розділення хвостатого ядра і лушпини неповне – вони зв’язані між собою хвостато-сочевицеподібними сірими мостами (pontes grisei caudatolenticulares), які проходять між волокнами внутрішньої капсули і містять два вищезгаданих види нейронів. Ростровентрально, де кількість волокон внутрішньої капсули поступово зменшується, ядра з’єднуються між собою достатньо великим скупченням нервових клітин. Хвостате ядро та лушпина розвиваються з матриксу навколо бічних шлуночків і є похідними нової кори telencephalon. Ці ядра складають разом стріатум або неостріатум, філогенетично старші бліді кулі мають назву палідум або палеостріатум (paleostriatum). Бліді кулі, так само як і підталамічне ядро (n. subthalamicus) проміжного мозку, мають походження з ембріонального гангліозного горбка проміжного мозку, тому в деяких підручниках бліді кулі описують як окрему частину проміжного мозку – субталамуса.
Загалом утворення смугастого тіла належать до екстрапірамідної системи. Мигдалеподібне тіло звичайно розглядають як утворення нюхового мозку і складову частину лімбічної системи. Функції огорожі поки що остаточно не з’ясовані (вважають, що, подібно до мигдалеподібного тіла, вона не є частиною екстрапірамідної системи).
Хвостате ядро (nucleus caudatus) має вигляд видовженого та дугоподібно вигнутого навколо таламуса тіла. Його передній кінець, стовщений у вигляді головки (caput), визначається в товщі лобової частки півкулі; донизу вона досягає передньої пронизаної речовини; присередньо головка виступає в порожнину переднього рогу бічного шлуночка. Середня частина ядра зветься тілом (corpus); вона розміщена в тім’яній частці півкулі нa дні центральної частини бічного шлуночка.
Задній кінець ядра – хвіст (cauda), поступово потоншуючись, іде по верхній стінці нижнього рогу і досягає мигдалеподібного тіла. Назовні від хвостатого ядра розташований досить товстий прошарок білої речовини – внутрішня капсула (capsula interna), яка відокремлює його від сочевицеподібного ядра.
Сочевицеподібне ядро (nucleus lentiformis) повністю міститься в товщі білої речовини і має як на фронтальних, так і на горизонтальних зрізах трикутну форму. Дві тонкі білі пластинки поділяють це ядро на лушпину та бліді кулі. Бічна мозкова пластинка (lamina medullaris lateralis) відмежовує лушпину від блідих куль; присередня мозкова пластинка (lamina medullaris medialis) відмежовує присередню бліду кулю (globus pallidus medialis) від бічної блідої кулі (globus pallidus lateralis). Назовні від сочевицеподібного ядра лежить прошарок білої речовини – зовнішня капсула (capsula externa), яка відокремлює його від огорожі.
Огорожа (claustrum) має вигляд вертикальної пластинки сірої речовини, витягнутої в ростральнокаудальному та дорсовентральному напрямку. На її латеральній поверхні є пальцеподібні вдавлення, які утворились у місцях розгалуження гачкоподібного пучка (fasciculus uncinatus). Суміжними структурами огорожі по латеральній поверхні є острівцева та скронева кора, від якої вона відокремлена крайньою (найбільш зовнішньою) капсулою (capsula extrema), а по медіальній – лушпина та, дещо вентральніше, мигдалеподібне тіло.
Огорожа належить до найменш вивчених структур мозку. За мікроструктурою ядро відрізняється від підкоркових утворень поліморфізмом клітинного складу, а від кори – відсутністю пошарового розташування його структур. За величиною та характером функціональних зв’язків (наявністю проекційних зон різних модальностей) огорожа нагадує проекційні зони кори, в яких, однак, відсутня виражена соматотопія. Функціональна структура огорожі особливо незвичайна: від базальних ядер вона відрізняється топографічною визначеністю аферентних (сомаестезичного, акустичного, візуального, нюхового, інтероцептивного) та еферентних представництв, а від кори – відсутністю чіткого розмежування цих представництв і величиною зон їх взаємного перекриття. Досліди з прямою електричною стимуляцією огорожі свідчать про її участь в організації всіх видів еферентної діяльності (орієнтовній, харчовій, оборонній, емоційній) і в підтриманні загального рівня збудливості (бадьорості та сну). Спеціальні досліди показали, що огорожа приймає участь і у вищій нервовій діяльності. Таким чином, огорожа являє собою своєрідну (в морфологічному та фізіологічному аспектах) структуру мозку, в якій поєднуються властивості і функції, властиві і кортикальним, і субкортикальним утворенням головного мозку.
Мигдалеподібне тіло (corpus amygdaloideum) – досить велике утворення, розміщене у скроневій частці півкулі спереду від нижнього рогу бічного шлуночка. Нині мигдалеподібне тіло розглядають як проміжне утворення між мозковою корою та ядрами. Воно багате на пептидергічні нейрони і складається з комплексу ядер та навколомигдалеподібної кори (cortex periamygdaloideus).
До ядер мигдалеподібного тіла відносяться: поверхнево розташоване кіркове ядро мигдалика (nucleus amygdale corticalis); центральне ядро мигдалика (nucleus amygdale centralis); основне ядро (nucleus basalis), яке поділяється на дрібноклітинне присереднє основне ядро мигдалика (nucleus amygdale basalis medialis) i на крупноклітинне бічне основне ядро мигдалика (nucleus amygdale basalis lateralis); бічне ядро мигдалика (nucleus amygdale lateralis); присереднє ядро мигдалика (nucleus amygdale medialis).
Всі ядра мигдалеподібного тіла поділяють на дві групи:
1) філогенетично старішу кортикомедіальну групу (до неї належать кортикальні та центральні ядра);
2) філогенетично молодшу базолатеральну групу (до неї відносяться базальні та латеральні ядра).
Електрофізіологічні дослідження вказують, що тільки ядра кортикомедіальної групи одержують нюхові імпульси; між тим, ядра базолатеральної групи одержують зорові та слухові імпульси.
Електрична стимуляція ядер мигдалеподібного тіла та їх оточення викликає різноманітні вегетативні та емоційні реакції. Зокрема, при стимуляції мигдалеподібного тіла під час нейрохірургічних операцій з приводу важких епілепсій у людей з’являлось відчуття люті і страху або відчуття заспокоєння та зменшення напруги. Причому реакція залежала від емоційного стану людей на момент стимуляції.
Значним досягненням останніх років, яке пояснює функціональні зв’язки базальних ядер, є нейрохімічне вивчення дофамінергічних нейрональних систем. Одна з таких систем починається нейронами чорної речовини середнього мозку, аксони яких йдуть через ніжку мозку, внутрішню капсулу, бліді кулі – до холінергічних нейронів хвостатого ядра і здійснюють на них гальмівний вплив. Дегенерація цього нігростріатного дофамінергічного шляху призводить до розгальмування гальмівного впливу хвостатого ядра на paleostriatum і визнається головним чинником паркінсонізму (основні симптоми якого: збіднілість рухової активності; підвищення тонусу м’язів; тремтіння пальців рук, нижньої щелепи, голови).
Важливим шляхом дофамінергічної системи є також мезолімбічний шлях, який починається в середньому мозку від міжніжкового ядра (nucleus interpeduncularis) і зв’язує смугасте тіло зі структурами лімбічної системи і корою лобової частки півкуль великого мозку. Цей шлях бере участь у контролі за настроєм і поведінкою, а також контролює початок рухового акту та рухів афективної реакції (наприклад, рухів, що супроводжують емоції).
Ці дані розкривають зв’язки та інтегративні механізми смугастого тіла, визначаючи його не тільки як просто рефлекторно-автоматизований центр рухової системи (екстрапірамідний центр), а ще і як утворення, що організує зосередження уваги та емоцій на одному, найважливішому в цей момент руховому акті, що досягається шляхом одночасного гальмування усіх інших процесів, які можуть перешкодити його здійсненню.
Бічні шлуночки
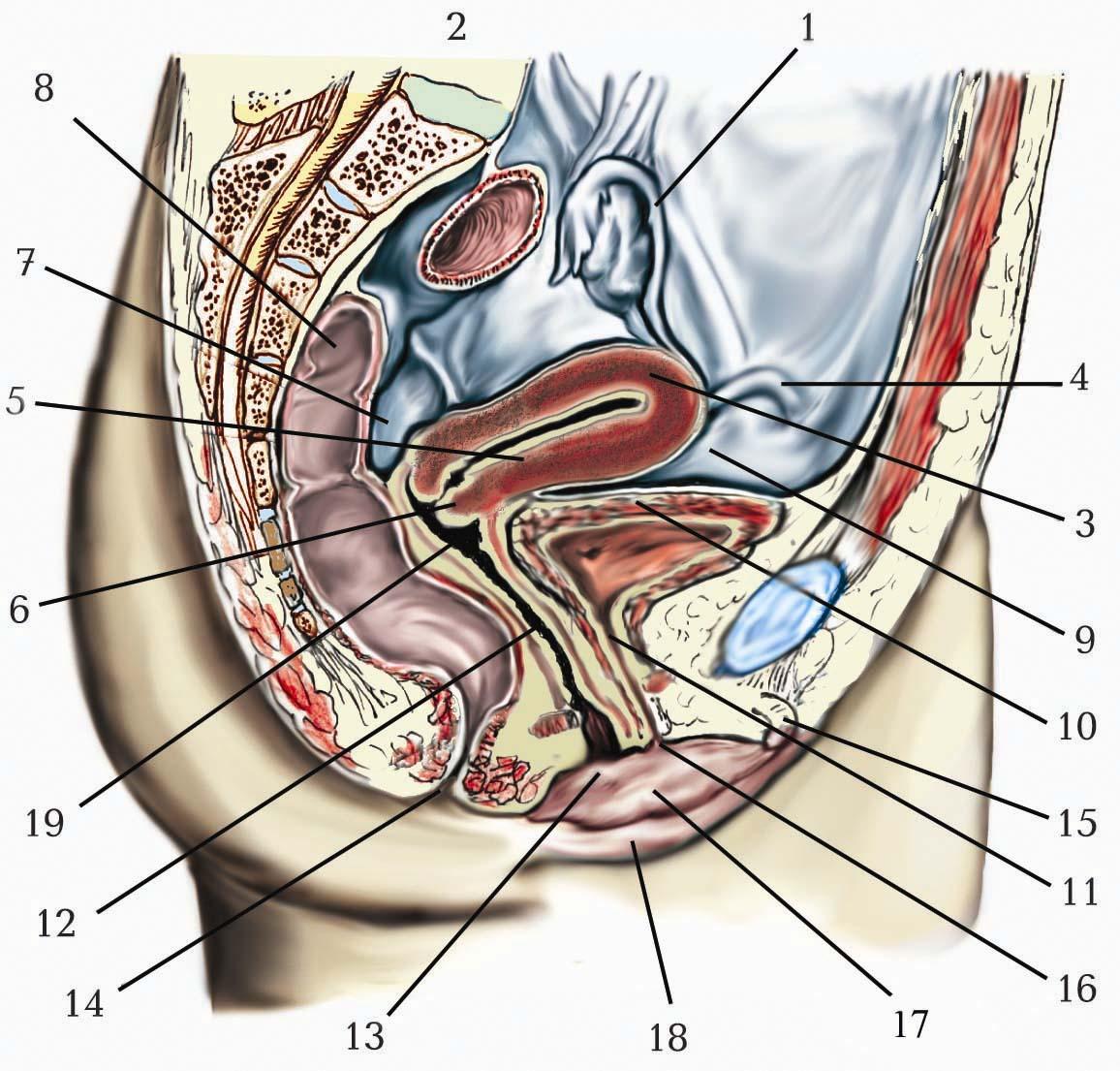
Бічний шлуночок (ventriculus lateralis) являє собою парну щілиноподібну порожнину, яка розміщена в глибині півкуль у горизонтальній площині.
Розрізняють правий і лівий бічні шлуночки, причому умовились вважати лівий шлуночок першим, правий – другим.
Кожний бічний шлуночок вистелений епендимою, містить спинномозкову рідину та сполучається з ІІІ шлуночком за допомогою міжшлуночкового отвору, отвору Монро (foramen interventriculare, foramen Monro). Складовими частинами бічного шлуночка є порожнини всіх часток мозкової півкулі, за винятком острівцевої та лімбічної. У кожному бічному шлуночку розрізняють: лобовий, потиличний, скроневий роги та центральну частину.
Лобовий ріг (передній ріг), cornu frontale (cornu anterius) бічного шлуночка залягає всередині лобової частки. Верхньою, передньою і частково нижньою стінками порожнини лобового рогу є передні відділи мозолистого тіла. Нижньою і бічною стінками є головка хвостатого ядра, яка випинає в порожнину переднього рогу. Присередньою стінкою переднього рогу є тонка прозора перегородка. Задньої стінки передній ріг не має, тому що в верхніх відділах порожнина його безпосередньо переходить у порожнину центральної частини, а в глибині його задніх відділів залягає міжшлуночковий отвір.
Центральна частина (pars centralis) бічного шлуночка є порожниною тім’яної частки півкулі і становить сплощену згори донизу вузьку щілину. Верхньою стінкою порожнини є стовбур мозолистого тіла; нижньою – таламус, тіло хвостатого ядра, погранична смуга; присередньою – тіло склепіння. Таламус в порожнині центральної частини прикритий рядом утворень. Присередньо на ньому лежить потужне судинне сплетення (plexus choroideus), під яким тягнеться тіло склепіння. Назовні від судинного сплетення, будучи почасти ним прикрита, простягається вузька і тоненька пластинка мозкової речовини, прикріплена нижньою поверхнею до таламуса – прикріплена пластинка (lamina affixa). За походженням в ембріогенезі прикріплена пластинка є стінкою першого мозкового міхура, який прикріплюється в цьому місці до верхньої стінки другого міхура. Назовні від цієї пластинки залягає вузька погранична смуга (stria terminalis), яка являє собою пучок мієлінових нервових волокон, що починаються від мигдалеподібного тіла і відділяють таламус від хвостатого ядра. Ззаду центральна частина безпосередньо переходить у потиличний ріг.
Потиличний ріг (задній ріг), cornu occipitale (cornu posterius), є порожниною потиличної частки. Вона дуже вузька і майже вся виповнена вдавленим з присереднього її боку утворенням – пташиною острогою (calcar avis), яка відповідає заглибленню мозкової кори на присередній поверхні півкулі – остроговій борозні. Трохи догори і наперед від пташиної остроги залягає невелика складка (часто слабко виявлена) – цибулина заднього рогу (bulbus cornus posterior). Пташина острога та цибулина заднього рогу утворюють присередню стінку потиличного рогу. Згори і зовні порожнина потиличного рогу оточена волокнами мозолистого тіла, які одержують тут, а також у скроневому розі, назву покриву (tapetum). Нижню стінку потиличного рогу (яка продовжується в нижню стінку скроневого рогу) утворює трикутної форми площа – обхідний трикутник (trigonum collaterale), на якій іноді відзначається невелике випинання – обхідне підвищення (eminentia collateralis).
Скроневий ріг (нижній ріг), cornu temporale (cornu inferius), розміщений у товщі скроневої частки півкулі. Його порожнина ближче до присередньої периферії частки становить трохи зігнуту донизу, наперед і досередини щілину. Верхня стінка скроневого рогу утворена хвостом хвостатого ядра, нижня – обхідним підвищенням, бічна – покривом мозолистого тіла. Присередня стінка скроневого рогу утворена аммоновим рогом, який дуже випинається в порожнину бічного шлуночка. Випинання аммонового рогу бере початок в ділянці центральної частини бічного шлуночка, утворює згин у напрямі до потиличного рогу і, спускаючись згори вниз і ззаду наперед, дугоподібно продовжується у скроневий ріг, утворюючи його присередню стінку. Присередніше від аммонового рогу розміщена біла вузька торочка морського коника, яка зростається з ним і продовжується наперед до ноги морського коника, а ззаду переходить у ніжку склепіння.
В центральній частині та у скроневому розі бічного шлуночка міститься судинне сплетення бічного шлуночка, яке є бічним краєм судинного сплетення ІІІ шлуночка і з’єднується з ним крізь міжшлуночковий отвір. Судинне сплетення бічного шлуночка є похідним м’якої оболони головного мозку і проникає в порожнину шлуночка крізь судинну щілину (fissura choroidea), яка розміщена в його центральній частині між таламусом та склепінням, а в нижньому розі – між пограничною смугою (stria terminalis) та торочкою морського коника. Судинне сплетення бічного шлуночка має потовщення в місці переходу центральної частини шлуночка у скроневий ріг, яке лежить на trigonum collaterale i зветься судинним клубком (glomus choroideum).
Біла речовина кінцевого мозку
Весь простір між сірою речовиною мозкової кори та сірою речовиною базальних ядер займає біла речовина. Вона складається з великої кількості нервових волокон, які йдуть у різних напрямках і утворюють провідні шляхи кінцевого мозку. Нервові волокна поділяють на: 1) асоціативні; 2) комісуральні; 3) проекційні .
Асоціативні волокна кінцевого мозку (fibrae associationes telencephali) складають основу білої речовини, з’єднують різні ділянки кори мозку в межах однієї півкулі і забезпечують можливість для кори виконувати асоціативні та інтегративні функції. Наявність численних зв’язків між окремими кірковими ділянками є можливим поясненням того факту, що втрата мозкової функції внаслідок вогнищевого пошкодження кори не завжди постійна. Можливо, деякі волокна залишаються інтактними і здатні поновлювати частину втрачених функцій після певних вправ протягом деякого часу.
Серед асоціативних волокон розрізняють короткі та довгі волокна.
Короткі асоціативні волокна (fibrae associationes breves) дугоподібно з’єднують сусідні звивини і звуться дугоподібними волокнами великого мозку (fibrae arcuatae cerebri).
Довгі асоціативні волокна (fibrae associationes longae) з’єднують між собою звичайно кору різних часток однієї півкулі. Верхній поздовжній пучок (fasciculus longitudinalis superior) розташований у верхній частині півкулі. Він з’єднує кору лобової, тім’яної, потиличної та скроневої часток між собою. Лобово-скронева частина цього пучка огинає задній край sulcus lateralis i має назву – дугоподібний пучок (fasciculus arcuatus). Вважають, що дугоподібний пучок з’єднує скроневі (центр Верніке) і лобові (центр Брока) мовні центри. Нижній поздовжній пучок (fasciculus longitudinalis inferior) зв’язує кору скроневої та потиличної часток між собою. Гачкуватий пучок (fasciculus uncinatus) з’єднує кору лобового полюса та нижніх відділів лобової частки з корою передніх відділів скроневої частки. Пояс (cingulum) являє собою один з асоціативних пучків лімбічної системи, огинає мозолисте тіло на всьому його протязі і з’єднує підмозолисте поле (area subcallosa) з корою всіх структур лімбічної частки до гачка включно. Нижній потилично-лобовий пучок (fasciculus occipitofrontalis inferior) розповсюджується від лобової до потиличної частки, проходячи через вентральну частину capsula externa.
У півкулях проходять також інші довгі асоціативні волокна, найважливішими серед яких є прямовисні потиличні пучки (fasciculi occipitales verticales) та верхній потилично-лобовий пучок (fasciculus occipitofrontalis superior).
Комісуральні нервові волокна кінцевого мозку (fibrae commissurales telencephali) з’єднують симетричні ділянки кори обох мозкових півкуль і утворюють волокна мозолистого тіла (fibrae corporis callosi), спайку склепіння (commissura fornicis) i передню спайку (commissura anterior) (commissura posterior, commissura habenularum являють собою структури проміжного мозку).
Проекційні нервові волокна з’єднують кору кінцевого мозку з нижчерозташованими відділами ЦНС до спинного мозку включно. Проекційні волокна в білій речовині півкулі ближче до кори утворюють віялоподібне скупчення – променистий вінець (corona radiata), головна частина волокон якого сходиться у внутрішню капсулу.
Внутрішня капсула (capsula interna) є ростральним продовженням основи ніжки мозку відповідного боку, тому на фронтальних розрізах вона має вигляд косої смужки, нервові волокна якої віялоподібно розгортаються і розподіляються досить товстим шаром між сочевицеподібним ядром, з одного боку, хвостатим ядром і таламусом – з іншого боку. На горизонтальному розрізі внутрішня капсула має вигляд відкритого тупого кута з його частинами: передньою ніжкою, коліном та задньою ніжкою.
Передня ніжка (crus anterius) розташована між хвостатим ядром та передньою половиною сочевицеподібного ядра. До складу проекційних волокон її входять: 1) лобово-мостовий шлях (tractus frontopontineus) – йде від мозкової кори лобової частки до ядер моста; 2) передня променистість таламуса (radiatio thalami anterior) – з’єднує мозкову кору лобової частки з таламусом).
Коліно внутрішньої капсули (genu capsulae internae) розміщене між передньою та задньою ніжками. Воно утворене кірково-ядерними волокнами (fibrae corticonuclearis) (входять до складу пірамідних шляхів і з’єднують кору передцентральної звивини з руховими ядрами черепних нервів).
Задня ніжка (crus posterius) розташована між таламусом та задньою половиною сочевицеподібного ядра. Вона має (за відношенням до сочевицеподібного ядра) три складові частини: таламо-сочевицеподібну, засочевицеподібну та підсочевицеподібну .
Безпосередньо до коліна внутрішньої капсули прилягає таламо-сочевицеподібна частина (pars thalamolentiformis), яку складають:
1) кірково-спинномозкові волокна (fibrae corticospinales) – входять до складу пірамідних шляхів, з’єднують кору передцентральної звивини та рухові ядра передніх рогів спинного мозку;
2) кірково-червоноядерні волокна (fibrae corticorubrales) – зв’язують кору лобової частки з червоним ядром;
3) кірково-сітчасті волокна (fibrae corticoreticulares) – йдуть від передцентральної та зацентральної звивини до ретикулярної формації;
4) кірково-таламічні волокна (fibrae corticothalamicae) – зв’язують мозкову кору з ядрами таламуса;
5) таламо-тім’яні волокна (fibrae thalamoparietales) – частина волокон висхідних шляхів кіркового напрямку;
6) центральна таламічна променистість (radiatio thalami centralis) – більша частина волокон висхідних шляхів кіркового напрямку, які прямують до мозкової кори передцентральної та зацентральної звивини.
Позаду сочевицеподібного ядра розміщена засочевицеподібна частина (pars retrolentiformis) задньої ніжки внутрішньої капсули, яка складається із:
1) задньої таламічної променистості (radiatio thalamica posterior) – волокна її йдуть від бічного колінчастого тіла та подушки таламуса до кори острогової борозни;
2) потилично-мостових волокон (fibrae occipitopontinae) – з’єднують мозкову кору потиличної частки з ядрами моста;
3) потилично-покрівельних волокон (fibrae occipitotectales) – йдуть від мозкової кори потиличної частки до lamina tecti середнього мозку.
Під сочевицеподібним ядром розташована підсочевицеподібна частина (pars sublentiformis) задньої ніжки внутрішньої капсули, яка містить:
1) кірково-покрівельні волокна (fibrae corticotectales);
2) скроне-мостові волокна (fibrae temporopontinae) – йдуть від кори скроневої частки до ядер моста;
3) кірково-таламічні волокна (fibrae corticothalamicae);
4) зорову променистість (коліно-острогові волокна), radiatio орtica (fibrae geniculocalcarinae) – волокна йдуть від бічного колінчастого тіла до кори острогової борозни;
5) слухову променистість (коліно-скроневі волокна), radiatio acustica (fibrae geniculotemporale) – волокна йдуть від присереднього колінчастого тіла до кори поперечних скроневих звивин.
Оболони головного мозку
Головний мозок вкритий з усіх боків такими ж самими трьома оболонами мозку (meninges), як і спинний:
1) черепна тверда оболона (тверда оболона головного мозку), dura mater cranialis (dura mater encephali);
2) черепна павутинна оболона (павутинна оболона головного мозку), arachnoidea mater cranialis (arachnoidea mater encephali);
3) черепна м’яка оболона (м’яка оболона головного мозку), pia mater cranialis (pia mater encephali). Дві останні об’єднуються під назвою м’яка оболона (leptomeninx) на противагу твердій оболоні (pachimeninx).
Тверда оболона головного мозку (dura mater encephali) складається з двох шарів щільної сполучної тканини. Зовнішній шар є окістям і щільно прилягає до кісток черепа. Внутрішній шар є власне менінгеальним шаром і повернений до дуже вузького підтвердооболонного простору. Внутрішній менінгеальний шар твердої оболони місцями відходить від її зовнішнього шару, утворюючи венозні пазухи (синуси). В ділянці зовнішнього краю великого потиличного отвору шари твердої оболони повністю розділяються. Зовнішній продовжується у вигляді окістя кісток черепа, а менінгеальний утворює “твердооболонний мішок” спинного мозку.
Від внутрішньої поверхні твердої оболони головного мозку відходить кілька відростків, які заходять у вигляді пластинок у щілини між окремими частинами головного мозку і відокремлюють їх одну від одної: серп великого мозку, серп мозочка, намет мозочка, діафрагма сідла .
Серп великого мозку (falx cerebri) нагадує за формою лезо серпа і розміщений прямовисно між обома півкулями великого мозку в fissura longitudinalis cerebri. У передній частині головного мозку серп своїм увігнутим краєм заглиблюється в щілину між лобовими частками, але не досягає мозолистого тіла, дозаду ж він стає вищим. Серп великого мозку починається спереду від crista galli решітчастої кістки і своїм опуклим краєм прикріплюється до боків сагітального шва на склепінні черепа, доходячи до protuberantia occipitalis interna, де попереду від нього з’єднується з верхньою поверхнею мозочкового намету.
Серп мозочка (falx cerebelli) має вигляд короткого і невисокго відростка, який заходить в задню вирізку мозочка між його півкулями і прикріплюється до crista occipitalis interna. Біля заднього краю потиличного отвору він закінчується двома ніжками, які обмежують отвір ззаду; його верхній опуклий край з’єднується з наметом мозочка.
Намет мозочка (tentorium cerebelli) проникає у fissura transversa cerebri i відокремлює потиличні частки великого мозку від мозочка. Намет мозочка має вигляд горизонтальної пластинки, опуклий задньолатеральний край якої прирощений до країв поперечних борозен os occipitale i до верхнього краю піраміди скроневої кістки. Передній вільний край намету увігнутий і має вирізку намету (incisura tentorii), яка (разом зі спинкою турецького сідла) обмежує отвір, через який проходить міст і середній мозок.
Діафрагма сідла (diaphragma sellae) закриває згори ямку турецького сідла, завдяки чому утворюється порожнина, яка містить гіпофіз. Всередині діафрагми є отвір, через який до гіпофіза проходить лійка (infundibulum).
В ділянці impressio trigemini на верхівці піраміди скроневої кістки тверда оболона головного мозку розщеплюється на два листки і утворює трійчасту порожнину (cavum trigeminale), яка містить чутливий вузол трійчастого нерва.
Тверда оболона головного мозку відокремлюється від павутинної оболони, що лежить під нею, щілиноподібною порожниною – підтвердооболонним простором (spatium subdurale), в якому є невелика кількість рідини. Павутинна оболона головного мозку не фіксована до твердої, за винятком зон вздовж венозних пазух, до яких вона прикріплена за допомогою павутинних зернистостей (granulationes arachnoideae).
Оскільки в підтвердооболонному просторі завжди міститься невелика кількість прозорої рідини (яка не має жодного відношення до спинномозкової рідини), павутинна оболона здатна зміщуватися відносно твердої без всякого тертя (це дає можливість для певних коливних зміщень півкуль головного мозку в порожнині черепа без пошкодження мозкової тканини).
Деякі сучасні анатоми (D. E. Haines, 1991; F. Van Denabeele et al., 1996), базуючись на даних електронної мікроскопії, підкреслюють, що хоча термін “підтвердооболонний простір, spatium subdurale” та термін “надтвердооболонний або позатвердооболонний простір, spatium epidurale seu extradurale” досить поширені у неврологічній літературі, проте за нормальних умов павутинна оболона прикріплюється до твердої оболони, а та – до черепа; при цьому, зрозуміло, не виникає порожнин або просторів. Поява зазначених просторів – наслідок механічного пошкодження або патологічного процесу, що штучно відокремлює павутинну оболону від твердої, а тверду – від черепа.
Павутинна оболона головного мозку (arachnoidea mater encephali) облягає мозок з усіх боків і вступає в зв’язок з м’якою оболоною, що лежить під нею, за допомогою численних волокон і перекладок, переплетення яких нагадує павутиння. Від м’якої оболони вона відокремлена підпавутинним простором (spatium subarachnoideum), заповненим спинномозковою рідиною. В ділянках частин мозку, що виступають (наприклад, над звивинами), павутинна та м’яка оболони тісно прилягають одна до одної; над заглибинами вони розходяться, бо м’яка оболона точно повторює рельєф мозку, а павутинна перекидається з одного виступу на інший. Таким чином, підпавутинний простір не являє собою суцільну порожнину однакової глибини, а складається з великої кількості з’єднаних між собою щілин, деякі з них досягають значних розмірів і називаються підпавутинними цистернами (cisternae subarachnoideae).
Найбільшою підпавутинною цистерною є задня мозочково-мозкова цистерна (велика цистерна), cisterna cerebellomedularis posterior (cisterna magna), яка розміщена у заглибині між мозочком та довгастим мозком. Вона сполучається з ІV шлуночком через його серединний отвір. Цю цистерну використовують для проведення підпотиличної пункції (для одержання спинномозкової рідини або для введення повітря в шлуночкову систему при пневмоенцефалографії); прокол при цьому здійснюють через задню атлантопотиличну перетинку .
Важливим з клінічної точки зору є те, що судини і нерви, які проходять через підпавутинний простір, розташовані у спинномозковій рідині. Тому при інфікуванні цього простору вони втягуються у запальний процес.
Вентрально задня мозочково-мозкова цистерна вільно сполучається з парною мосто-мозочковою цистерною (cisterna pontocerebellaris). Ця цистерна розташована у мосто-мозочковому куті і сполучається з ІV шлуночком через його бічний отвір. Ця цистерна, крім інших структур, містить хребтову артерію.
На базальній поверхні головного мозку послідовно розміщені:
1) між ніжками мозку – міжніжкова цистерна (cisterna іnterpeduncularis);
2) перед зоровим перехрестям – цистерна перехрестя (cisterna chiasmatis). Ця цистерна перехрестя містить в собі внутрішньочерепні частини внутрішніх сонних артерій. Коли внутрішня сонна артерія розгалужується на передню та середню мозкові артерії, підпавутинний простір розширюється і утворює, відповідно, навколомозолисту цистерну (cisterna pericallosa) за ходом передньої мозкової артерії і цистерну бічної ямки великого мозку (cisterna fossae lateralis cerebri) за ходом середньої мозкової артерії.
Навколомозолиста цистерна є непарною і продовжується по зовнішній поверхні мозолистого тіла аж до його валика. Там вона з’єднується з відносно великою чотиригорбковою цистерною (цистерною великої вени великого мозку), cisterna quаdrigeminalis (cisterna venae magnae cerebri). У цій цистерні, крім vena magna cerebri (вени Галена), міститься шишкоподібна залоза. Чотиригорбкова цистерна сполучається з задньою мозочково-мозковою цистерною і, крім того, з міжніжковою цистерною через оточуючу цистерну (cisterna ambiens), яка розміщена збоку від ніжки мозку.
Практичне значення має те, що в порожнині черепа і при виході з черепа черепних нервів оболони головного мозку супроводжують їх на невеликій відстані. Особливо важливо це для зорового нерва. Тверда оболона вистеляє зоровий канал. Коли вона досягає очної ямки, її зовнішній шар стає окістям ямки, а менінгеальний шар іде на всьому протязі зорового нерва і прикріплюється до склери. Всередині цього “твердооболонного мішка” розташований “підпавутинний мішок”, а також підтвердооболонний та підпавутинний простори, які є безпосереднім продовженням відповідних внутрішньочерепних просторів. Внаслідок таких анатомічних особливостей формуються шляхи розповсюдження крововиливів, інфекції, пухлин з порожнини черепа до ока і навпаки.
М’яка оболона головного мозку (pia mater encephali) утворена сполучною тканиною, багатою на нерви і кровоносні судини, має велике значення в живленні мозку. М’яка оболона проходить в усі заглибини та борозни мозку і зростається з ним за допомогою ектодермальної мембрани, утвореної крайовими астроцитами. М’яка оболона оточує судини, що проникають в мозок, утворюючи навколо них периваскулярну піальну мембрану, яка, зі зменшенням калібру судини, змінюється на периваскулярну межову гліальну мембрану, утворену астроцитами.
У певних місцях м’яка оболона головного мозку утворює складки (дуплікатури), які, зростаючись зі стоншеною до епендимного шару стінкою шлуночків мозку, становлять судинний прошарок (tela choroidea). Там, де в зазначеній складці є особливо багато кровоносних судин, утворюється судинне сплетення (plexus choroideus).
Шляхи циркуляції спинномозкової рідини
Шлуночкова система головного мозку складається з двох бічних шлуночків, які через міжшлуночкові отвори сполучаються з непарним третім шлуночком. Третій шлуночок через водопровід середнього мозку з’єднується з четвертим шлуночком. Четвертий шлуночок має сполучення з підпавутинним простором через серединний і бічні отвори. Кожний з чотирьох шлуночків має судинне сплетення (plexus choroideus). Найбільшим є сплетення бічного шлуночка.
Стінки шлуночків вкриті епендимними клітинами, які в певних ділянках мають структурно-функціональні особливості. До таких клітин, зокрема, належать епендимні клітини, що утворюють органи циркумвентрикулярної системи, і хороїдні епендимоцити.
У нижчих тварин доведена секреторна та рецепторна функції епендими. Розвиток шляхів циркуляції спинномозкової рідини у ссавців призвів до появи спеціалізованих структур, які об’єднують у циркумвентрикулярну систему . До складу цієї системи у людини, зокрема, входять невеличкі утворення епендимної глії, розміщені в ІІІ шлуночку – судинний орган lamina terminalis i підсклепінний орган (organum subfornicale); a також розташоване на дні ІV шлуночка біля входу в центральний канал зазаднє поле (area postrema). Циркумвентрикулярна система регулює тиск спинномозкової рідини і її хімічний склад, а також має потужні зв’язки з нейроендокринною системою гіпоталамуса. До цієї системи відносять також шишкоподібну залозу і гіпофіз, участь яких у регуляції ліквороутворення обговорюється.
У тварин до циркумвентрикулярної системи входять також підспайковий орган і парафіз, які у людини є транзиторними органами, виникають в ембріональному періоді та існують протягом невеликого часу.
Хороїдні епендимоцити вкривають м’яку оболону головного мозку, що впинається у просвіт шлуночків головного мозку і утворює судинні сплетення. Епендимоцити судинних сплетень входять до складу гемато-лікворного бар’єру (бар’єру між кров’ю та спинномозковою рідиною), крізь який відбувається ультрафільтрація крові з утворенням спинномозкової рідини (біля 500 мл на добу). При ультрафільтрації крові утворюється 70–90 % спинномозкової рідини; 10–30 % її продукують тканини ЦНС і виділяють через епендиму, розташовану поза межами судинних сплетень.
Спинномозкова рідина циркулює в шлуночках головного мозку, в центральному каналі спинного мозку і в підпавутинному просторі; її загальний об’єм у дороcлих складає біля 140 мл. Вона повністю поновлюється кожні 4–7 годин і за складом відрізняється від плазми крові низьким вмістом білка та підвищеною концентрацією натрію, калію та хлору. Спинномозкова рідина містить окремі лімфоцити (не більше 5 клітин на 1 мл); її тиск у людини, яка лежить, дорівнює 70–120 мм вод. ст.
Спинномозкова рідина виконує ряд функцій:
1) захисна (амортизація ударів і струсів мозку);
2) утворення гідростатичної оболони навколо мозку, його корінців і судин (завдяки чому зменшується натяжіння корінців і судин);
3) утворення оптимального рідкого середовища, що оточує структури ЦНС (підтримання постійного іонного балансу, який забезпечує нормальну діяльність нейронів і глії);
4) виведення метаболітів, що утворюються в мозковій тканині;
5) інтегративна (перенесення гормонів та інших біологічно активних речовин).
Простір, в якому міститься спинномозкова рідина, є замкненим. Всмоктування компонентів спинномозкової рідини в кров відбувається в ділянках павутинних зернистостей; у незначній мірі воно здійснюється епендимою судинних сплетень. Павутинні зернистості (granulationes arachnoideae), через які спинномозкова рідина переходить шляхом фільтрації з підпавутинного простору в венозні пазухи твердої оболони та вени губчатки (vv. diploicae), являють собою утворення павутинної оболони головного мозку і мають вигляд бахромчастих відростків різної величини. Вони прилягають до твердої оболони головного мозку в місцях розташування sinus sagittalis superior, sinus transversus і коло інших венозних пазух.
Павутинні зернистості вдавлюються у тверду оболону, потоншуючи її, і разом з нею вдавлюються у венозні пазухи або у внутрішню поверхню черепних кісток (в місцях розміщення в них кров’яних озер – розширень венозних судин кісток черепа). На місцях цих вдавлень під впливом павутинних зернистостей кістка потоншується і утворюються ямочки зернистості (foveolae granulares). В ділянках павутинних зернистостей, що занурюються у венозні пазухи і кров’яні озерця, спинномозкова рідина відділяється від крові тільки шаром гліальних клітин і судинним ендотелієм, що створює структурні передумови для її зворотної фільтрації у кров.