Функціональні системи ЦНС та їх структурне забезпечення
Пірамідна рухова система
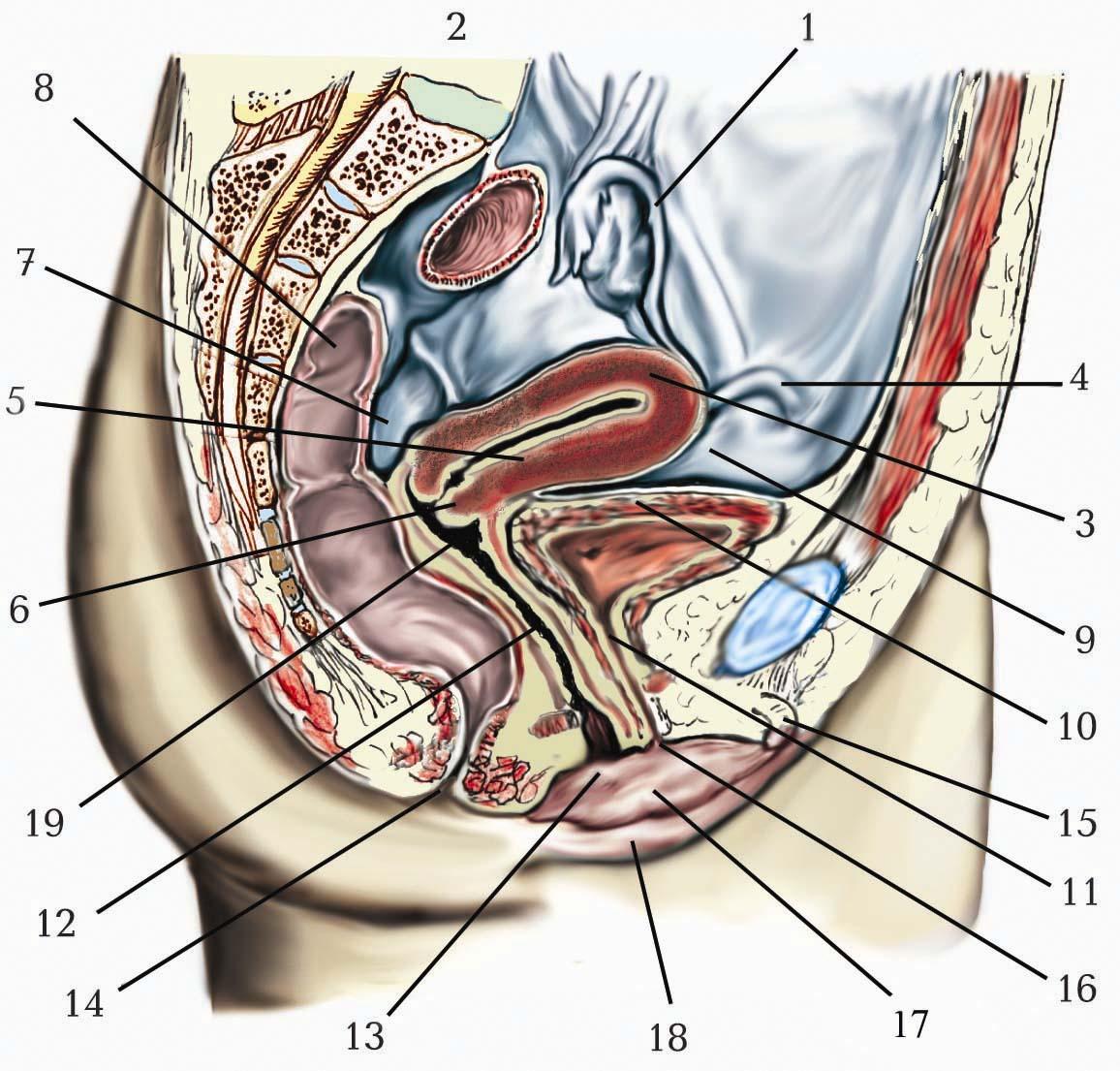
Шляхами, які проводять імпульси цілеспрямованих вольових рухів, є кірково-спинномозковий (tractus corticospinalis) і кірково-ядерний (tractus corticonuclearis) пірамідні шляхи. Волокна цих шляхів є аксонами нейронів, розташованих переважно в передцентральній звивині (цитоархітектонічних полях 4 та 6а), в задній третині верхньої та середньої лобових звивин (поле 6 а), в середній третині лобових звивин (поле 8), у верхній частині зацентральної звивини (поля 1, 2, 3).
Цитоархітектонічне поле представлене вузькою смужкою кори, що розповсюджується вздовж центральної борозни до верхнього краю півкулі і далі по передній частині прицентральної часточки на присередній поверхні півкулі. Нейрони, які забезпечують іннервацію глотки і гортані, розташовані в його нижній частині, поблизу від бічної борозни. Потім у висхідному порядку йдуть нейрони до м’язів лиця, руки, тулуба і ноги (“руховий гомункулус”). Цитоархітектонічне поле 6а розташоване попереду поля 4 також у межах передцентральної звивини.
Особливістю цитоархітектонічної організації поля 4 є потужний розвиток п’ятого шару, який містить гігантські пірамідні клітини Беца. Ще з класичних робіт Лоренто де Но, надрукованих у 1943 році, відомо, що пірамідні нейрони розташовані нерівномірно, групами з вертикальними зв’язками між нейронами ІІ і ІV шарів моторної кори. Підтвердження групування нейронів кори в радіально орієнтовані колонки було отримано в електрофізіологічних дослідженнях В. Маунткасла у шестидесяті роки. Пізніше низкою досліджень було встановлено, що вертикально орієнтовані колонки є елементарними функціональними комірками рухової кори. Кожна вертикальна колонка – структурний модуль – це група клітин, де гігантська пірамідна клітина займає центральне положення, а навколо неї – 2–3 великі або середні пірамідні клітини, дендрити яких ідуть щільним пучком. Між такими клітинними ансамблями (мікроколонками) спостерігається взаємопроникнення дендритів, що полегшує синаптичні контакти набору мікроколонок з одним і тим же аферентним волокном, і таким чином з одного аферентного волокна може залучатися до реакції ціла система мікроколонок.
Вважають, що рухові колонки здатні збуджувати або гальмувати групу функціонально однорідних мотонейронів. У той же час виявлено, що нейрони кори, які регулюють діяльність якого-небудь м’яза, не сконцентровані у межах тільки однієї колонки. Показано, що для колонок характерна тонка функціональна спеціалізація. Наприклад, існують окремі колонки моторної кори, пов’язані зі швидкими (фазними, чи фазичними) та повільними (тонічними) рухами. Вважають, що кожна колонка керує суглобом, тобто від колонки йдуть команди до різних м’язів, діяльність яких має відношення до даного суглоба. Встановлено, що вертикальні колонки спеціалізуються і щодо положення суглоба – збудження одних колонок викликає згинання відповідного суглоба, а збудження інших – фіксацію цього суглоба. Іншими словами, рухова колонка значною мірою являє собою функціональне об’єднання нейронів, які регулюють діяльність декількох м’язів, що діють на той чи інший суглоб. Таким чином, у колонках пірамідних нейронів моторної кори представлені не стільки м’язи, скільки рухи.
Поряд з пірамідною системою у моторній корі починається екстрапірамідна система. Ця система відрізняється тим, що аксони її кіркових клітин ідуть до базальних ядер, мозочка, червоного ядра, присінкових ядер, ретикулярної формації та інших структур стовбура мозку, котрі по tr. rubrospinalis, fibrae reticulospinales, fibrae olivospinales та інших низхідних шляхах впливають на спинномозкові нейрони. В цілому, пірамідні та екстрапірамідні шляхи – це єдиний механізм, завдяки якому виконується складний цілеспрямований рух при збереженні рівноваги та орієнтації у просторі.
При ураженні моторної кори у людини спостерігається параліч, або плегія, тобто повна відсутність можливості виконання цілеспрямованих довільних рухів. Особливо помітно це проявляється у кистях рук, стопах, мімічних м’язах, пов’язаних з артикуляцією. В окремих випадках розвивається парез, тобто часткове зменшення об’єму та сили рухів. При односторонніх ураженнях моторної кори відбувається поступове відновлення рухових функцій. При двосторонніх ураженнях воно, як правило, відсутнє (це вказує на значення двосторонніх зв’язків між моторною корою обох півкуль для компенсації рухових порушень). Проте за рахунок інтеграції пірамідної системи з екстрапірамідною системою навіть при двосторонньому ураженні рухової кори у ряді випадків можливе часткове відновлення деяких видів довільних рухів (головним чином таких, що не потребують точної та тонкої координації).
У моторну кору приходить аферентація від тактильних, пропріоцептивних, вестибулярних, вісцеральних, зорових та слухових рецепторів, як безпосередньо від відповідних проекційних, так і від асоціативних ділянок кори. Важливо підкреслити, що через систему асоціативних волокон рухова ділянка кори пов’язана з багатьма зонами протилежної півкулі, що має велике значення для ефективного керування довільними рухами.
Особливе значення для діяльності моторної кори має еферентація, яка поступає до неї від рецепторів м’язів, сухожилків та суглобів. Вхід до моторної кори від пропріоцептивних м’язів топічно специфічний. Це забезпечує реалізацію рефлексів, які замикаються через пірамідні клітини безпосередньо на альфа-мотонейронах спинного мозку. Така організація лежить в основі високого ступеня самоорганізації та самовдосконалення руху шляхом корекції управління рухами на основі сигналів зворотного зв’язку.
Як відомо, основними асоціативними зонами є лобова асоціативна ділянка (поля 8, 9, 10, 11, 12) та тім’яна асоціативна ділянка (поля 5, 7, 39, 40). Ці зони мають пряме відношення до формування задуму руху та організації самого руху. Зокрема, показано, що тім’яна кора, яка отримує аферентні імпульси від проекційних ділянок кори, а також від асоціативних ядер таламуса, має велике число еферентних виходів до моторної кори, що полегшує формування команди невимушеної дії на базі аферентного синтезу. Лобова асоціативна ділянка (поля 8, 9, 10, 11, 12), приймаючи участь в реалізації психічних процесів, одночасно є місцем організації цілеспрямованої діяльності, в тому числі за рахунок прийняття рішення та формування програми дії. Ця ділянка у людини має також безпосереднє відношення до формування усної мови (центр Брока, поля 44 і 45, сумісно з полями 6 та 8) та письмової мови (поле 8 з полями 39 та 40 тім’яної асоціативної ділянки кори).
Інформація про задум руху передається відповідним центрам моторної кори, котрі направляють рухові команди до альфа-мотонейронів спинного мозку (по пірамідних шляхах), а також до базальних ядер, мозочка, червоного ядра, присінкових ядер та ретикулярної формації стовбура мозку (по екстрапірамідних шляхах).
Пошкодження окремих ділянок асоціативних областей призводить до порушення складних форм довільної цілеспрямованої дії, тобто до апраксії, у тому ж числі до ідеаторної апраксії, чи апраксії задуму (порушення послідовності рухів, необхідних для виконання задачі, а також нездатність за усним завданням у повному обсязі виконати завдання; поля 39 та 40), моторної апраксії (порушення виконання складного завдання, у тому ж числі і за наказом та за наслідуванням; поле 40) і конструктивної апраксії (у порушенні здатності конструювати ціле з частин; поле 39). Крім того, порушення асоціативних ділянок може призводити до розвитку моторної афазії (поле 8 при участі полів 6, 44 і 45), аграфії (поля 8 і 40), а також до порушення інших тонких рухів, що потребують участі вищих відділів мозку.
Архіпалеокортекс, котрий включає структури стародавньої і давньої кори великого мозку, являє собою одну з важливих інтегративних систем мозку та є неспецифічним активатором усіх видів кіркової діяльності. Окрім інших функцій (нюх, увага, емоції, регуляція вегетативних процесів), архіопалеокортекс забезпечує здійснення біологічно важливих вроджених рефлексів (зокрема, пошуковий, харчовий, статевий, захисний) та їх рухових компонентів. Можливо, що вказані структури архіпалеокортекса містять вроджені рухові програми, реалізація яких забезпечує прояв вказаних рефлексів.
Поле 4 дає біля 40 % волокон пірамідних шляхів, які забезпечують окремі точні цілеспрямовані рухи. Вони починаються від нейронів п’ятого шару кори, причому лише 3,4–4 % волокон починаються від гігантських пірамідних клітин Беца; ці волокна є швидкопровідними і мають товсту мієлінову оболонку. Решта волокон пірамідних шляхів починається від малих нейронів п’ятого шару кори (60 % з них є мієлінізованими і 40 % немієлінізованими). Стимуляція поля 4 індукує загальні рухи окремих м’язів, у той час коли стимуляція поля 6 викликає складні рухи, такі як рухи всієї руки чи ноги.
Волокна кірково-спинномозкового шляху починаються головним чином від нейронів кори верхніх двох третин передцентральної звивини, проходять послідовно через передній відділ задньої ніжки внутрішньої капсули, середню частину основи ніжки мозку, передню частину моста, збираючись компактно у вигляді пірамід довгастого мозку. 90 % волокон цього шляху перехрещуються на межі зі спинним мозком, утворюючи перехрестя пірамід, і далі йдуть у складі бічних канатиків спинного мозку протилежного боку у вигляді бічного кірково-спинномозкового шляху (tractus corticospinalis lateralis). Близько 10 % волокон продовжуються в передніх канатиках спинного мозку свого боку у вигляді переднього кірково-спинномозкового шляху (tractus corticospinalis anterior) і перехрещуються посегментно. Передній кірково-спинномозковий шлях є дуже варіабельним: може бути асиметричним або взагалі відсутнім. Найчастіше він не поширюється нижче грудного відділу спинного мозку.
Приблизно 90 % волокон кірково-спинномозкового шляху перемикаються в проміжній зоні між задніми і передніми рогами на вставних нейронах (інтернейронах), в тому числі і на гальмівних клітинах Реншоу, які в свою чергу утворюють синапси з альфаі гамма-мотонейронами передніх рогів спинного мозку. Це має велике значення, бо, наприклад, згинання в суглобі неможливе без одночасного скорочення м’язів-згиначів і розслаблення м’язів-розгиначів. Біля 10 % волокон прямує безпосередньо до мотонейронів передніх рогів; це волокна, що забезпечують іннервацію м’язів дистальних відділів кінцівок.
Аксони мотонейронів передніх рогів спинного мозку в складі передніх корінців і гілок спинномозкових нервів досягають скелетних м’язів шиї, тулуба і кінцівок, забезпечуючи їх свідомі рухи. Причому завдяки тому, що всі волокна кірково-спинномозкового шляху є перехрещеними, однобічне ураження головного мозку або половини спинного мозку призводить до паралічу м’язів протилежної половини тіла нижче місця ураження.
Волокна кірково-ядерного шляху (tractus corticonuclearis) починаються головним чином від кори нижньої третини передцентральної звивини, проходять через коліно внутрішньої капсули і досягають розташованих у стовбурі головного мозку нейронів рухових ядер черепних нервів свого та протилежного боку. Винятком з цього є ядро VІ пари, нижня частина рухового ядра VІІ пари та ядра ХІІ пари черепних нервів, волокна до яких йдуть тільки від кори протилежної півкулі. Аксони нейронів рухових ядер черепних нервів у складі гілок цих нервів прямують до скелетних м’язів голови та шиї і забезпечують їх двобічну (від лівої та правої півкуль) центральну іннервацію. Винятком є тільки мімічні м’язи нижньої половини лиця та м’язи язика, а також бічний прямий м’яз очного яблука, які мають однобічну гетеролатеральну центральну іннервацію. Є також дані про те, що волокна ядра ІV пари залишаються неперехрещеними.
Як окрему частину пірамідних шляхів описують кірково-середньомозкові волокна (fibrae corticomesencephalicae), які починаються від очної ділянки 8 цитоархітектонічного поля (частина кори середньої лобової звивини спереду від поля 6а) і йдуть до ядер окорухових нервів (забезпечують співдружній поворот очей у протилежний бік).
Згідно з сучасними уявленнями, пірамідні шляхи не є тільки винятково руховими шляхами, але містять також різні низхідні регулюючі системи. Зокрема волокна, які йдуть від тім’яної частки, закінчуються на nucleus gracilis та nucleus cuneatus довгастого мозку, а також на substantia gelatinosa задніх рогів спинного мозку. Встановлено, що вони регулюють потужність чутливих імпульсів, які генерують ці ядра.
Екстрапірамідна рухова система
Крім вищезгаданих рухових центрів і пірамідних шляхів, є багато інших центрів і шляхів, які забезпечують рухову активність; ці утворення дістали назву екстрапірамідних. Екстрапірамідна система філогенетично старіша за пірамідну і об’єднує ядра і шляхи, що автоматично (несвідомо) регулюють роботу м’язів і забезпечують підтримку м’язового тонусу та координацію рухів. Крім того, екстрапірамідна система регулює налаштування і готовність до дії рухового апарату, забезпечує перерозподіл тонусу м’язів під час рухів, створення фону для здійснення швидких, точних, диференційованих рухів, реалізацію як безумовно-рефлекторних захисних і співдружніх рухів, так і завчених, стереотипних, автоматизованих (у тому числі професійних навичок), надаючи їм плавності і завершеності. Зараз вивчаються зв’язки екстрапірамідної системи з лімбічною системою. Визначена її участь в інтегративних механізмах вищої нервової діяльності (особливо в механізмах емоційно-афективних реакцій організму), встановлена наявність тісного взаємозв’язку розвитку кінетичних та психічних функцій.
Треба відзначити зв’язки екстрапірамідної і пірамідної систем. Як уже описувалось нами раніше, кірково-мостові волокна від лобової, тім’яної, скроневої, потиличної часток півкуль великого мозку проходять через внутрішню капсулу, йдуть у міст і утворюють синапси з нейронами його ядер, nuclei pontеs. Ці нейрони посилають свої аксони в кору протилежної півкулі мозочка. Внаслідок цих зв’язків кора мозочка одержує, так би мовити, копію всіх рухових імпульсів, що йдуть від кори півкуль великого мозку. Мозочок отримує також (по висхідних шляхах мозочкового напрямку) інформацію про всю рухову активність на периферії. Таким чином мозочок стає здатний контролювати та координувати вольові рухи через екстрапірамідну систему.
Мозочок входить до складу декількох функціональних нейронних кіл екстрапірамідної системи. До таких ланцюгів зворотного зв’язку належить так званий трикутник Гюлейна – Молларета – від червоного ядра до нижньооливного комплексу через tractus tegmentalis centralis (його частину tractus rubroolivaris); від мозочка до мозочкової кори, до зубчастого ядра; і знову – до червоного ядра.
Мозочок, який є частиною комплексного регуляторного механізму із зворотним зв’язком, діє в якості координаційного центру по забезпеченню рівноваги тіла і підтриманню м’язового тонусу.
Archicerebellum (див. розділ “Мозочок”) отримує інформацію про положення голови у просторі від рецепторів вестибулярної системи (волоскових клітин плями маточки та плями мішечка) і про рух голови – від рецепторів півколових проток (волоскових клітин ампульних гребенів) – через кінестетичні імпульси, які йдуть від присінкових ядер VIII пари черепних нервів через нижні ніжки мозочка. Це дозволяє мозочку постійно відповідно моделювати рухові імпульси спинного мозку, що забезпечує збереження рівноваги незалежно від положення тіла і його руху.
Paleocerebellum отримує аферентні імпульси зі спинного мозку по передньому та задньому спинномозково-мозочкових шляхах і від довгастого мозку через клино-мозочкові волокна. Еферентні імпульси від paleocerebellum модулюють активність м’язів, що забезпечують антигравітацію, і забезпечують м’язовий тонус, необхідний для підтримання рівноваги під час стояння чи руху. Спинномозкові імпульси мають соматотопічну проекцію на кору paleocerebellum, у кожній півкулі мозочка представлена гомолатеральна частина тіла. Кора прилеглих до черв’яка зон проектується на кулясте та коркоподібне ядра, кора черв’яка – на ядро вершини.
Еферентні волокна від нейронів ядер мозочка перехрещуються у верхніх мозочкових ніжках і досягають контрлатеральних червоних ядер. Низхідні червоноядерно-спинномозковий та червоноядерно-ядерний шляхи знову перехрещуються у decussatio tegmentalis anterior і регулюють активність рухових нейронів стовбура мозку і спинного мозку, розташованих гомолатерально відносно ядер мозочка і гетеролатерально відносно червоних ядер. Імпульси від мозочкових ядер надходять до неспецифічних (серединних та інтраламінарних) ядер таламуса і від них – до хвостатого ядра та лушпини (стріатум або неостріатум), впливаючи таким чином і на екстрапірамідну систему. Співдружня дія archicerebellum i paleocerebellum забезпечує регуляцію м’язового тонусу і тонку координовану роботу м’язів – синергістів і антагоністів, які забезпечують нормальну ходу і стояння.
Neocerebellum отримує аферентну імпульсацію від кори півкуль великого мозку, в основному від полів Бродмана 4 і 6, по кірково-мосто-мозочкових шляхах. Він також отримує значну кількість аферентації від розміщеного в довгастому мозку нижнього оливного комплексу по оливо-мозочковому шляху, а цей комплекс у свою чергу отримує імпульси від розміщеного в середньому мозку червоного ядра по центральному покривному шляху (tractus tegmentalis centralis). У мозочок надходить інформація про кожний довільний рух, що планується, попередньо. Ці рухи модифікуються і коректуються шляхом інгібування пірамідних та екстрапірамідних рухових імпульсів по зубчасто-таламічно-кіркових шляхах, які йдуть від пов’язаного з корою neocerebellum зубчастого ядра мозочка і закінчуються в тій же частині кори великого мозку, де зароджуються рухові імпульси. Екстрапірамідна система доповнює пірамідну систему свідомих, вольових рухів, піднімаючи її функціонування на вищий рівень, при якому кожний довільний рух стає дуже точно підготовленим та здійснюється плавно і скоординовано. Пірамідні шляхи (через інтернейрони) і екстрапірамідний ланцюг нейронів врешті-решт зустрічаються на мотонейронах переднього рогу спинного мозку, здійснюючи на них частково активуючий і частково гальмуючий вплив.
Як уже підкреслювалося в розділі “Внутрішня будова спинного мозку”, серед мотонейронів ЦНС розрізняють альфа-мотонейрони, які діють на екстрафузальні м’язові волокна в складі нейромоторних одиниць, та гамма-мотонейрони, які діють на інтрафузальні м’язові волокна в складі пропріорецепторів – м’язових веретен (регулюють їх чутливість і забезпечують оптимізацію їх роботи).
Нейромоторні одиниці в різних м’язах мають певні відмінності, які, зокрема, визначаються іннерваційним індексом – відношенням числа мотонейронів (у нормі – це один мотонейрон), аксони яких закінчуються на зовнішній мембрані м’язового волокна, до числа м’язових волокон. У м’язів, які виконують точні швидкі рухи, іннерваційний індекс малий; у потужних м’язів кінцівок – великий. Наприклад, окорухові м’язи мають індекс 1 : 3, а двоголовий м’яз плеча – 1 : 100 (тобто один мотонейрон діє своїм нейротрансмітером на сто м’язових волокон). Внаслідок цього, дія поодинокої одиниці м’яза кінцівки – одне швидке скорочення, яке виникає під впливом одного імпульсу, що викликає виділення нейротрансмітера із закінчення одного мотонейрона, – відповідно виявляється грубою.
Важливим інтегративним центром екстрапірамідної системи є також таламус. Таламус пов’язаний з корою півкуль великого мозку значною кількістю аферентних і еферентних волокон. Аферентні імпульси з таламуса надходять до кори великого мозку і смугастого тіла; зі смугастого тіла волокна йдуть у чорну речовину, з якої основна частина волокон спрямовується в ретикулярну формацію і невелика частина – в бічну бліду кулю. Бічна бліда куля через сочевицеподібну петлю (ansa lentiformis) пов’язана з ретикулярною формацією, із субталамічним ядром. Присередня бліда куля тісно пов’язана аферентними волокнами з бічною і посилає еферентні волокна до вентролатеральних ядер таламуса. Останні віддають аферентні волокна в кору великого мозку. Таким чином ядра екстрапірамідної системи об’єднуються у функціональні системи (нейронні кола), в яких спостерігається постійна імпульсація (зі швидкістю 10 імпульсів на секунду), забезпечуючи контроль реакцій за типом зворотних зв’язків:
1) таламус – стріопалідарні ядра – таламус;
2) таламус – кора великого мозку – стріопалідарні ядра – таламус;
3) таламус – кора великого мозку – міст – мозочок – таламус.
Як уже зазначалося раніше, в межах екстрапірамідної системи виділяють “стріопалідарну систему”. До стріарної її частини належить хвостате ядро і лушпина, а до палідарної – бліді кулі. Філогенетично молодша стріарна частина гальмує палідарну частину. Ураження цієї системи призводить до порушення м’язового тонусу і до рухових розладів. Якщо відбувається ураження стріарної частини, рухи м’язів розгальмовуються (що має місце при хореї); якщо відбувається ураження палідарної частини, рухи м’язів гальмуються (що має місце при паркінсонізмі). На даний момент накопичилось багато відомостей про те, що рухова кора півкуль великого мозку керує рухом, використовуючи інформацію, що надходить як по сенсорних шляхах від інших відділів кори, так і від моторних програм, що генеруються в ЦНС у базальних ядрах і мозочку і доходять до рухової кори через таламус, де вони, можливо, попередньо взаємодіють. Імовірно, існує загальний біохімічний механізм актуалізації моторних програм через ріст активності дофамінергічних і норадренергічних систем у головному мозку. Ці системи визначають властивість гальмувати гальмівні інтернейрони і тим самим розгальмовувати нейронні мережі.
Є також дані про наявність у striatum ГАМКергічних гальмівних інтернейронів, що тонічно гальмують запрограмовані нейронні ланцюги, які керують позою і рухами, а самі гальмуються дофамінергічними нейронами чорної речовини. Доведено, що норадренергічні волокна проектуються в мозочок, їх активація також призводить до гальмування гальмівних нейронів. Зараз багато дослідників поділяють точку зору, що дофамінергічні та норадренергічні шляхи беруть участь у гальмуванні гальмівних інтернейронів у структурах, які роблять внесок в ініціацію і руховий контроль поведінки. Таким чином, у базальних ядрах мозочку вже закладений механізм, що може активувати рухові програми, які там зберігаються, або загальмувати їх. За відкриття ролі дофамінергічних нейронів чорної речовини в процесах регуляції рухової активності шведський дослідник Арвід Карлссон одержав у 2000 році Нобелівську премію.
Встановлено, що при хворобі Паркінсона руйнування чорної речовини патологічним процесом супроводжується зменшенням продукції дофаміну, який виробляється клітинами її компактної частини і надходить по нігростріальних шляхах у хвостате ядро. У хвостатому ядрі дофамін пригнічує активність його нейронів, виконуючи функцію “гальмування гальмування”. При нестачі дофаміну гальмівна функція хвостатого ядра за рахунок підвищення в його власних нейронах кількості ацетилхоліну посилюється, що призводить до гіпокінезії.
Серед усіх шляхів ЦНС найбільшою є кількість екстрапірамідних шляхів. Головним екстрапірамідним шляхом стовбура головного мозку є центральний покривний шлях (tractus tegmentalis centralis) (описаний в розділі “Середній мозок”). У білій речовині спинного мозку проходять волокна таких екстрапірамідних шляхів:
1) tractus vestibulospinalis lateralis et medialis;
2) tractus rubrospinalis;
3) fibrae olivospinales;
4) tractus tectospinalis;
5) fibrae reticulospinales, які поділяються на tractus pontoreticulospinalis, tractus bulboreticulospinalis, tractus caerulospinalis;
6) tractus solitariospinalis та інші (функція цих шляхів описана в розділі “Спинний мозок”).
Довгі аферентні провідні шляхи
Довгі аферентні провідні шляхи передають інформацію від рецепторів і призначені для:
1) забезпечення свідомої чутливості (соматичної та вісцеральної);
2) проведення імпульсів до сенсорних і моторних центрів у стовбурі мозку, мозочку, проміжному мозку, базальних ядрах (несвідома чутливість);
3) забезпечення зворотного зв’язку для здійснення рефлексів (соматичних та вісцеральних).
У цьому розділі ми зупинимося на соматичних шляхах. Розділення аферентних шляхів на соматичні шляхи свідомої та соматичні шляхи несвідомої чутливості, на лемніскові та екстралемніскові тощо, дещо умовне, позаяк сенсорні функції – це інтегральний наслідок діяльності різних анатомічно і функціонально взаємопов’язаних і взаємозалежних систем і утворень. Разом з тим для систематизації аферентних шляхів і для зручності викладання ми послідовно зупинимося на:
1) соматосенсорних шляхах свідомої чутливості (лемніскових і екстралемніскових);
2) соматосенсорних шляхах несвідомої чутливості. Крім того, в складі соматосенсорних шляхів свідомої чутливості ми розглянемо окремо: 1) шляхи епікритичної чутливості; 2) шляхи протопатичної чутливості.
Соматосенсорні шляхи свідомої чутливості
Соматосенсорні шляхи свідомої чутливості (аферентні шляхи кіркового напрямку) традиційно поділяють на лемніскові й екстралемніскові. З часу появи робіт W. Magоun (1952) i П. K. Анохіна (1958) під терміном “лемніскова система” розуміють всю систему від рецептора до відповідних ділянок кори лобових і тім’яних часток. До лемніскових шляхів відносять ті шляхи, волокна яких беруть участь в утворенні lemniscus medialis, це:
1) tractus gangliobulbothalamocorticalis;
2) tractus gangliospinothalamocorticalis;
3) tractus ganglionucleothalamocorticalis.
Порушення чутливості, пов’язані з патологією систем і утворень, анатомічно розташованих поза межами згаданих вище аферентних шляхів, були названі екстралемнісковими. До екстралемніскових шляхів свідомої чутливості багато авторів відносять неспецифічні структури лімбічної системи, спинномозково-сітчастий шлях (tractus spinoreticularis), який зараз трактують як філогенетично старішу палеоспіноталамічну частину tractus gangliospinothalamocorticalis. Стовбурово-діенцефальні відділи ретикулярної формації, до яких прямує tractus spinoreticularis, нині розглядають як частину лімбічної системи, “серцем” якої є морський коник. Морський коник, до якого йдуть імпульси екстралемніскових шляхів, функціонує як мультисенсорна полімодальна структура великого мозку.
Кожний соматосенсорний шлях свідомої чутливості йде від периферичного рецептора до кори великого мозку. Шлях починається рецептором шкіри або скелетно-м’язової системи. Деякі чутливі модальності характеризуються специфічними рецепторами і мають дискретні чутливі шляхи, але як периферичні рецептори, так і центральні шляхи проявляють тенденцію частково перекривати інші модальності (різні тонкі різновиди больової, температурної, тактильної, пропріоцептивної чутливості).
Тіло першого нейрона всіх соматосенсорних шляхів розташоване в чутливих вузлах спинномозкових або черепних нервів. Тіло ІІ нейрона розташоване в спинному мозку або в мозковому стовбурі. Його аксони здійснюють перехрест (або в передній білій спайці спинного мозку, або в довгастому мозку) і йдуть далі через стовбур, де всі шляхи об’єднуються в присередню петлю (lemniscus medialis), а потім утворюють синапси на тілі ІІІ нейрона в протилежному таламусі. Аксони ІІІ нейрона йдуть від таламуса через задню ніжку внутрішньої капсули до соматосенсорної ділянки кори великого мозку.
При аналізі у хворого втрати чутливості, проведення імпульсів якої здійснюється через той чи інший шлях, лікар повинен знати:
1) локалізацію рецептора;
2) шлях через периферійну нервову систему;
3) локалізацію тіла І, ІІ, ІІІ нейронів;
4) місце і рівень перехресту аксонів ІІ нейрона;
5) канатик спинного мозку (передній, бічний, задній), в складі якого проходить шлях;
6) шлях у стовбурі мозку;
7) таламічні транслюючі ядра;
8) білу речовину між таламусом і корою;
9) відповідне кіркове представництво.
Є певні морфологічні закони, які допомагають запам’ятати початок, напрямок, положення і закінчення спинномозкових шляхів:
1) закон периферичного розташування довгих волокон;
2) закон філогенетичного розшарування;
3) закон розшарування, викликаного рівнем входження нервових волокон або топографією тіла;
4) закон розділення чутливих шляхів чутливими модальностями.
Головним шляхом епікритичної чутливості, який відповідає за проведення імпульсів свідомої пропріоцептивної чутливості (суглобово-м’язова чутливість, стереогноз, вібраційна чутливість, відчуття тиску і маси) і тактильної чутливості (дотик) є tractus ganglio-bulbothalamocorticalis. Різновиди чутливості, за передачу імпульсів яких відповідає tractus gangliobulbothalamocorticalis, є дуже специфічними, що стосується стимулу, положення, просторових і часових відмінностей (дискримінації). Тому їх іноді називають дискримінативними модальностями. Можна узагальнити, що tractus gangliobulbothalamocorticalis відповідає за передачу екстероцептивних і пропріоцептивних імпульсів епікритичної чутливості. Цей шлях і його розгалуження повністю займають задній канатик спинного мозку, тому він ще має назву шляху заднього канатика.
Рецептори цього шляху локалізовані в шкірі та скелетно-м’язовій системі. Рецептори в шкірі включають тільця Фатера – Пачіні та Мейснера. Аксони нейронів, пов’язаних з рецепторами в шкірі, є середніми по товщині (ІІ альфата ІІ бета-волокна). Рецептори в скелетно-м’язовій системі – це м’язові веретена та сухожилкові органи Гольджі. Аксони нейронів, пов’язаних з цими рецепторами, є товщими (І альфата І бета-волокна).
Тіло першого нейрона цього шляху розташоване в чутливих вузлах спинномозкових нервів (псевдоуніполярні нейрони). Аксони проникають у спинний мозок у складі задніх корінців спинномозкових нервів і поділяються в задніх канатиках спинного мозку на висхідні та низхідні гілки.
Таким чином, задні канатики спинного мозку містять аферентні аксони первинних рецепторних або чутливих нейронів, клітинні тіла яких розташовані за межами ЦНС. Низхідні гілки аксонів задніх корінців ідуть каудально в задніх канатиках у вигляді міжпучкового пучка (fasciculus interfascicularis), який продовжується каудально в перегородково-крайовий пучок (fasciculus septоmarginalis). Волокна цих пучків формують у межах заднього канатика в шийному відділі – кому Шульца, в грудному – овальне поле Флексіга, в крижовому – трикутник Філіпе – Гомб (Gombault). Ці низхідні волокна закінчуються в пластинках І, ІV, V і частково VІ та VII (приблизно відповідає розташуванню ядер задніх та бічних рогів – nucleus proprius et nucleus dorsalis) i можуть впливати на висхідні потоки аферентної інформації.
Висхідні гілки аксонів задніх корінців формують в задньому канатику тонкий пучок (fasciculus gracilis) та клиноподібний пучок (fasciculus cuneatus), які прямують у гомолатеральному канатику відповідно до однойменних ядер довгастого мозку – nucleus gracilis et nucleus cuneatus. Частина висхідних волокон заднього канатика прямує до пов’язаного з мозочком додаткового клиноподібного ядра довгастого мозку (n. cuneatus accessorius) і об’єднується під загальною назвою – спинномозково-клиноподібні волокна (fibrae spinocuneatae).
Оскільки волокна збираються в задній канатик від куприкового до шийного відділу спинного мозку, то вони підпорядковуються закону розшарування за рахунок рівня входження і, таким чином, відображають топографію тіла. Присередньо розташований тонкий пучок одержує волокна від рецепторів нижньої кінцівки і нижньої частини тулуба через 19 нижніх спинномозкових сегментів. Прибічно розташований клиноподібний пучок одержує волокна від рецепторів верхньої кінцівки і верхньої половини тулуба через 12 верхніх спинномозкових сегментів, на рівні яких він тільки і визначається.
Тіло II нейрона tractus gangliobulbothalamocorticalis розташоване в nucleus gracilis i nucleus cuneatus довгастого мозку (одна з його назв – bulbus cerebri). Низхідні волокна цих ядер утворюють клино-спинномозкові і тонко-спинномозкові волокна (fibrae cuneospinales et fibrae gracilis spinales). Висхідні волокна цих ядер утворюють цибулинно-таламічний шлях (tractus bulbothalamicus), який перехрещується в перехресті присередніх петель (decussatio lemniscorum medialis) у дорсальній частині довгастого мозку. Волокна цибулинно-таламічного шляху до їх перехресту мають назву внутрішніх дугоподібних волокон (fibrae arcuatae internae). Цибулинно-таламічний шлях утворює основу присередньої петлі (lemniscus medialis), до якої потім приєднуються tractus spinothalamicus anterior, tractus spinothalamicus laterаlis, tractus nucleothalamicus.
Аксони нейронів тонкого та клиноподібного ядер після перехресту йдуть у складі присередньої петлі через стовбур мозку і досягають вентролатеральної групи ядер таламуса (тіло III нейрона). Аксони III нейронів проходять через задню ніжку внутрішньої капсули і закінчуються в корі передцентральної звивини, зацентральної звивини та верхньої тім’яної часточки.
Назву tractus gangliospinothalamocorticalis мають два анатомічно і функціонально різних шляхи протопатичної чутливості: 1) шлях, що відповідає за відчуття грубого дотику і тиску (входить до складу lemniscus medialis як передній спинномозковоталамічний шлях); 2) шлях больової і температурної чутливості (входить до складу lemniscus medialis як бічний спинномозково-таламічний шлях).
Тіло І нейрона tractus gangliospinothalamocorticalis – шляху, що відповідає за відчуття грубого дотику і тиску (різновиди тактильної чутливості), розміщене в чутливих вузлах спинномозкових нервів (псевдоуніполярні нейрони). Дендрити цих нейронів у складі спинномозкового нерва і його гілок прямують до рецепторів шкіри. Тактильні подразнення сприймаються чутливими тільцями Мейснера і дотиковими менісками (диски Меркеля), відчуття тиску – тільцями Фатера – Пачіні. Аксони І нейрона потрапляють через задні корінці в задній канатик, де у складі заднього власного пучка (fasciculus proprius posterior) піднімаються на 2–15 сегментів і утворюють синапси з нейронами задніх рогів (тіло II нейрона). В усіх закордонних підручниках з анатомії тілом II нейрона цього шляху вважають власне ядро (спинномозкові пластинки III та ІV), nucleus proprius (laminae spinales III et ІV). У підручнику анатомії М. Г. Привеса – драглисту речовину (substantia gelatinosa). Аксони II нейрона спускаються донизу на 1–2 сегменти (що треба враховувати при визначенні рівня ураження цього шляху у хворих) і перехрещуються в передній білій спайці. Далі вони утворюють передній спинномозково-таламічний шлях (tractus spinothalamicus anterior), який міститься на периферії переднього канатика спинного мозку. У стовбурі мозку аксони II нейрона входять до складу lemniscus medialis i разом з нею досягають нейронів вентролатеральної групи ядер таламуса (тіло III нейрона). Аксони III нейрона йдуть крізь задню ніжку внутрішньої капсули до кори зацентральної звивини.
Тіло І нейрона tractus gangliospinothalamocorticalis – шляху больової та температурної чутливості, представлене найменшими за розмірами псевдоуніполярними нейронами чутливих вузлів спинномозкових нервів. Дендрити псевдоуніполярних нейронів проходять у складі спинномозкових нервів і їх гілок, закінчуючись екстерорецепторами в шкірі: при цьому вільні нервові закінчення сприймають біль, тепло і холод, тільця Краузе – холод, тільця Руффіні (за деякими даними) – тепло. Аксони псевдоуніполярних клітин (волокна, які вони утворюють, відносяться до тонких мієлінових волокон групи III-А і майже безмієлінових волокон групи С) входять у спинний мозок через бічні відділи задніх корінців і, піднявшись у спинному мозку на 1–2 сегменти в задньобічному шляху бічного канатика (крайовий пояс Лісауера), tractus dorsolateralis, закінчуються безпосередньо або через інтернейрони задньобічного шляху на тілі II нейрона.
Зазначені інтернейрони мають важливе значення для регуляції (модуляції) потоків больових імпульсів. Встановлено, що збуджуючим нейромедіатором між аксонами нейронів чутливих вузлів спинномозкових нервів і перикаріонами II нейронів шляху болю та температури є речовина Р, і усунення больових відчуттів реалізується через рецептори опіоїдних пептидів, вбудованих в мембрану терміналі аксона чутливого нейрона (приклад феномена пресинаптичного гальмування). Джерелом опіоїдного пептиду енкефаліну є інтернейрон. Це багато в чому пояснює знеболюючі ефекти застосування препаратів морфію або, наприклад, голковколювання.
З приводу питання про тіло II нейрона цього шляху є давнє прикре непорозуміння. У більшості російських та українських підручників зазначено, що тілом II нейрона шляху больової та температурної чутливості є власне ядро (nucleus proprius); в усіх закордонних підручниках – драглиста речовина (спинномозкова пластинка II), substantia gelatinosa (lamina spinalis II). Драглиста речовина являє собою транслюючу (релейну) систему, в якій периферійні стимули модулюються допоміжними або інгібуючими впливами вищих надспинномозкових центрів. Останнє положення лягло в основу найбільш розповсюдженої в даний час так званої “ворітної теорії болю” (Р. Мелзак, 1981).
Аксони II нейрона перехрещуються в передній білій спайці і утворюють бічний спинномозково-таламічний шлях (tractus spinothalamicus lateralis), розміщений у бічних канатиках спинного мозку. Цей шлях іде в стовбурі мозку і в складі присередньої петлі досягає вентролатеральної групи ядер таламуса (тіло III нейрона). Аксони III нейрона цього шляху прямують до кори зацентральної звивини.
До лемніскових шляхів належить також шлях тактильної, больової, температурної та свідомої пропріоцептивної чутливості для голови та шиї tractus ganglionucleothalamocorticalis. Тіло І нейрона тієї частини цього шляху, яка відповідає за проведення імпульсів тактильної, больової та температурної чутливості, розташоване в чутливому вузлі трійчастого нерва (ganglion trigeminale) (псевдоуніполярні нейрони). Аксони І нейрона у складі чутливого корінця трійчастого нерва, radix sensoria, потрапляють в міст і прямують до чутливих ядер трійчастого нерва (тіло II нейрона): 1) nucleus principalis (ядро тактильної чутливості); 2) nucleus spinalis (ядро больової та температурної чутливості). Nucleus mesencephalicus трійчастого нерва (відповідає за свідому пропріоцептивну чутливість) побудоване з псевдоуніполярних нейронів, дендрити яких проходять через ganglion trigeminale без перерви, транзитом. Це єдиний випадок у нервовій системі людини, коли чутливі клітини (тіло І нейрона) розташовані в межах ЦНС. Таким чином, n. mesencephalicus можна розглядати як чутливий вузол, що “перемістився” у стовбур мозку.
- spinalis є видовженим, тому аксони І нейрона, що прямують до нього, утворюють спинномозковий шлях трійчастого нерва (tractus spinalis nervi trigemini).
- mesencephalicus також є видовженим, тому дендрити його псевдоуніполярних нейронів утворюють середньомозковий шлях трійчастого нерва (tractus mesencephalicus nervi trigemini).
Аксони чутливих ядер трійчастого нерва утворюють tractus nucleothalamicus, перехрещуються на протязі стовбура мозку і закінчуються у вентролатеральній групі ядер таламуса (тіло III нейрона для більшості волокон).
Аксони III нейронів (II нейронів для свідомої пропріоцептивної чутливості) проходять крізь задню ніжку внутрішньої капсули і прямують до нижньої третини передцентральної та зацентральної звивин.
Описані вище соматосенсорні шляхи свідомої чутливості відносяться до лемніскової системи і мають таку загальну особливість, що аксони їх II нейронів (n. mesencephalicus n. trigemini треба розглядати як виняток) утворюють у стовбурі мозку петлі, які всі з’єднуються в присередню петлю, lemniscus medialis. Таким чином, присередня петля складається з tractus bulbothalamicus, спинномозкової петлі (яка утворена tractus spinothalamicus anterior et tractus spinothalamicus lateralis) i трійчастої петлі (утвореної tractus nucleothalamicus).
Крім лемніскових шляхів, є також екстралемніскові шляхи свідомої чутливості. До них у першу чергу відноситься розташований в бічному канатику спинного мозку спинномозково-сітчастий шлях (tractus spinoreticularis). Спинномозково-сітчастий шлях (відповідно до найбільш розповсюдженої точки зору він є філогенетично старшим і може бути названий палеоспіноталамічним шляхом) утворений більшою частиною волокон tractus spinothalamicus lateralis. Так само як і tractus spinothalamicus lateralis, він відноситься до шляху протопатичної чутливості. Вважають, що він передає імпульси дифузного болю. Дифузний біль відрізняється від локального, гострого болю (останній передається через tractus spinothalamicus lateralis) тим, що є розлитим, доходить до свідомості людини після деякого прихованого періоду і відчувається довше від дії подразника. Тіло І, II, III нейронів шляху імпульсів гострого і дифузного болю співпадають; tractus spinoreticularis відрізняється від tractus spinothalamicus lateralis тільки тим, що є полісинаптичним і його колатералі утворюють численні синапси з нейронами ретикулярної (сітчастої) формації.
На сьогодні встановлено, що перерив tractus spinothalamicus lateralis, класичного шляху больової та температурної чутливості, призводить до щонайменше тимчасової втрати цих видів чутливості на гетеролатеральному (протилежному) боці тіла. Вісцеральна больова чутливість уражується при цьому в меншому ступені, бо вона, напевно, має більш дифузний шлях, ніж соматична чутливість.
Після хірургічної перерізки tractus spinothalamicus lateralis з метою полегшення болю (яка дістала назву трактотомії або кордотомії) рівень болю може через деякий час опуститися на декілька сегментів. Якщо рівень розрізу відповідає сегменту С2, то кінцевий рівень втрати чутливості з часом опускається до С5. У той же час частково больова (більшою мірою вісцеральна) і температурна чутливість може відновитися. Ці спостереження передбачають наявність цілої групи альтернативних шляхів. Дослідження на тваринах довели існування особливих, утворених маломієлінізованими або немієлінізованими волокнами, полісинаптичних гомолатеральних шляхів вісцеральної чутливості, які йдуть у сірій речовині спинного мозку. Мабуть, відгалуження подібних шляхів здатні формувати описаний в бічному канатику у людини спинномозково-шийний шлях, tractus spinocervicalis, який досягає ядер шийного відділу спинного мозку.
Соматосенсорні шляхи несвідомої чутливості
Переважна більшість усіх чутливих шляхів є шляхами зворотної аферентації, необхідної для здійснення будь-якого рефлексу. Свого часу шляхи до центрів екстрапірамідної системи розглядали саме в такому аспекті, але слід зазначити, що всі ці шляхи починаються від пропріорецепторів, тому безперечно можуть бути віднесені до шляхів несвідомої пропріоцептивної чутливості. Таким чином, всі соматосенсорні шляхи несвідомої чутливості можна поділити на:
1) пропріоцептивні шляхи мозочкового напрямку;
2) пропріоцептивні шляхи немозочкового напрямку.
Окремо слід відзначити розташований у бічному канатику спинномозково-покрівельний шлях (tractus spinotectalis), який йде до lamina tecti середнього мозку, проводить імпульси больової чутливості і забезпечує звуження зіниці, як реакцію на біль.
Тіло першого нейрона пропріоцептивних шляхів мозочкового напрямку розташоване в чутливих вузлах спинномозкових нервів. Аксони псевдоуніполярних аксонів через задні корінці спинномозкових нервів потрапляють у спинний мозок і утворюють:
1) волокна, що йдуть у складі бічного канатика до розташованої між переднім і заднім рогом проміжної зони – спинномозкової пластинки VII (lamina spinalis VII), де розміщене тіло II нейрона;
2) волокна, що йдуть у складі клиноподібного пучка заднього канатика до розташованого в довгастому мозку додаткового клиноподібного ядра (nucleus cuneatus accеssorius), де розміщене тіло II нейрона.
У відповідності до вищезгаданого, аксони II нейронів утворюють розташовані в бічному канатику спинного мозку задній та передній спинномозковомозочкові шляхи та розташований у довгастому мозку клино-мозочковий шлях (tractus cuneocerebellaris). Волокна останнього в межах довгастого мозку приєднуються до заднього спинномозково-мозочкового шляху і далі прямують у його складі.
Задній спинномозково-мозочковий шлях (tractus spinocerebellaris posterior) утворений аксонами тих нейронів спинномозкової пластинки VII, які формують заднє грудне (дорсальне ядро), nucleus thoracicus posterior (nucleus dorsalis). У цих нейронах, а також у нейронах гомологічного ядра довгастого мозку – додаткового клиноподібного ядра, перемикаються аксони задніх корінців С1–S5. Нейрони nucleus thoracicus posterior віддають свої аксони прибічно до периферії гомолатерального бічного канатика, де вони піднімаються догори як tractus spinоcerebellaris posterior. Оскільки nucleus thoracicus posterior не простягається нижче L2–L3, волокна від задніх корінців крижових і поперекових спинномозкових нервів піднімаються в бічних канатиках, для того щоб досягти рівня свого синапсування в nucleus thoracicus posterior. Подібним чином відбувається розподіл нервових волокон, які передають пропріоцептивні імпульси від верхніх кінцівок. Вони піднімаються в складі fasciculus cuneatus заднього канатика в довгастий мозок, де утворюють синапси на нейронах nucleus cuneatus accessorius. Нейрони nucleus cuneatus accessorius i nucleus thoracicus posterior віддають аксони, які через нижню мозочкову ніжку прямують до однойменної (гомолатеральної) частини мозочка.
У бічному канатику спинного мозку розміщений також передній спинномозково-мозочковий шлях (tractus spinocerebellaris anterior). Загалом його функція така сама, як і у заднього спинномозковомозочкового шляху, але він має і відмінності. Тіло II нейрона цього шляху розміщене у спинномозковій пластинці VII, але воно не настільки точно ідентифіковане, як nucleus thoracicus posterior для tractus spinothalamicus posterior. Tractus spinocerebellaris anterior проводить головним чином аксони, які починаються в каудальних відділах спинного мозку. Волокна tractus spinocerebellaris anterior двічі перехрещуються: 1) у передній білій спайці спинного мозку; 2) у верхньому мозковому парусі, до якого вони потрапляють через верхню мозочкову ніжку. Таким чином, tractus spinocerebellaris anterior, так само як і tractus spinocerebellaris posterior, з’єднує однойменні половини спинного мозку і мозочка. Обидва пропріоцептивні шляхи мозочкового напрямку підпорядковуються закону розшарування за рахунок рівня входження. Їх волокна поділяються соматотопічно таким чином: волокна, що передають імпульси від задніх корінців крижових спинномозкових нервів, розташовані дорcально, а від поперекових і грудних – вентрально.
Серед шляхів несвідомої пропріоцептивної чутливості немозочкового напрямку слід відзначити розташовані в бічному канатику спинного мозку:
1) спинномозково-оливний шлях (tractus spinoolivaris) – прямує до нижнього оливного комплексу довгастого мозку; продовженням його є оливо-мозочковий шлях (tractus olivocerebellaris).
2) спинномозково-присінковий шлях (tractus spinovestibularis) – прямує до присінкових ядер VIII пари черепних нервів; продовженням його є присінково-мозочковий шлях (tractus vestibulocerebellaris).
Нейроанатомічні механізми взаємодії та інтеграції відчуття і руху
Виходячи з матеріалу, що викладений у попередніх розділах підручника, кінцевий загальний руховий провідниковий шлях для всіх центрів (кіркових та підкіркових), залучених у рухову активність, закінчується аксоном альфа-мотонейрона, що іннервує скелетну мускулатуру, яка довільно скорочується. При цьому слід враховувати, що волокна рухових шляхів підходять не тільки безпосередньо до альфа-мотонейронів, але й до інтернейронів. У зв’язку з цим вони впливають на альфа-мотонейрони як безпосередньо, так і опосередковано, сповільнюючи або активуючи рефлекси між рецепторами та руховими нейронами. Таким чином, наприклад, передній ріг спинного мозку є не просто синаптичним центром, як уже підкреслювалось, а складним інтеграційним апаратом, який регулює рухову активність.
Ділянки кори великого мозку, які впливають на моторну систему через низхідні провідні шляхи, зв’язані одна з одною. Де ж джерело збудження, яке активує нейрони моторної кори? На це питання тепер, очевидно, можна відповісти таким чином: це збудження виникає в нейронах кори на найпізнішому етапі опрацювання всіх форм сенсорної інформації. На цьому етапі з’являються сигнали високого рівня абстракції, які відтворюють положення частин тіла в просторі та необхідність швидкого виконання тих чи інших рухів. Ця інформація, яка містить всі відомості про положення суглобів та напругу м’язів, слугує тією основою, керуючись якою моторна кора ініціює певні рухи.
Слід зазначити, що специфічні сенсорні системи, в загальних рисах подібно організовані, одночасно обробляють різні види чутливої інформації. Спеціалізовані рецепторні клітини, що розміщені в шкірі або в спеціальних органах чуття, сприймають фізичні або хімічні події, які відбуваються у зовнішньому світі. Потім ці події перетворюються у нейронну активність – у сигнали, які передаються нейронам, з’єднаним у впорядковану мережу, де і реалізується послідовне опрацювання інформації. Наприклад, різні аспекти зорових стимулів (такі як рух, колір та форма) послідовно переробляються у відокремлених, але паралельних каналах і в кінці кінців знову об’єднуються на рівні кори великого мозку. Інші форми сенсорної інформації піддаються подібній паралельній обробці.
Моторна система також здійснює послідовну обробку нервових сигналів. Вона діє за впорядкованим планом від ініціації руху корою до скорочення м’язів, які контролюють положення та стабільність суглобів, за командами мотонейронів. Паралельні модифікуючі системи мозочка, базальних ядер та інших центрів екстрапірамідної системи забезпечують координоване і плавне виконання рухової програми.
Число нервових елементів, яке використовує мозок для розрізнення властивостей подразника і для забезпечення швидкого, плавного і точного руху частин тіла, настільки велике, що це приховує від нас важливий висновок: різні сенсорні та моторні системи, можливо, в принципі анатомічно відокремлені та відносно незалежні одна від одної, але лише дуже невелика кількість сенсорних процесів завершуються, не приводячи до початку або уточнення рухів. Відчуття як кінцевий підсумок – досить рідкісне явище, окрім, мабуть, тих випадків, коли люди намагаються аналізувати свій власний чуттєвий досвід. Видатний російський фізіолог І. М. Сєченов у книзі: “Рефлексы головного мозга” більше ста років по тому писав: “Все бесконечное разнообразие внешних проявлений мозговой деятельности сводится окончательно к одному лишь явлению – мышечному движению. Смеется ли ребенок при виде игрушки, улыбается ли Гарибальди, когда его гонят за излишнюю любовь к родине, дрожит ли девушка при первой мысли о любви, создает ли Ньютон мировые законы и пишет их на бумаге – везде окончательным фактором является мышечное движение”.
І. П. Павлов розвинув та експериментально обґрунтував уявлення про аналізаторні системи, де кожний аналізатор – це певна анатомічно локалізована структура від периферичних рецепторних утворень до проекційних зон кори головного мозку. Він припустив, що, крім локальних проекційних зон кори, які виступають в якості “ядра кіркового кінця аналізатора” (або проекційної зони кори), існують периферичні зони представництва кожного аналізатора, так звані “зони розсіяних елементів”. В силу такої структурної організації всі аналізатори, включаючи і руховий аналізатор, своїми периферичними (кірковими) зонами перекриваються і утворюють вторинні проекційні зони кори, які І. П. Павлов уже тоді розглядав як асоціативні центри мозку, що складають основу для динамічної взаємодії всіх аналізаторних систем.
Відкриття І. П. Павловим аналізаторів і створення вчення про умовні рефлекси, в основі якого лежав об’єктивний аналіз динаміки нервових процесів, послужило основою для розвитку сучасних уявлень про динамічну локалізацію мозкових функцій – цілісне й одночасно диференційоване втягнення мозку в будь-яку форму його діяльності.
Інтегративно-пускові системи, або руховий аналізатор (за термінологією І. П. Павлова), посідає особливе місце в функціональній організації головного мозку. Це пов’язано з тим, що рухові ділянки стоять на виході інтегруючої і координуючої діяльності мозку і виконують функції запуску і контролю рухової діяльності, реалізації поведінкових актів. Сприйняття, адекватний вплив, надійне розпізнавання і висока здатність до диференціації подразника є необхідною передумовою діяльності рухових систем інтегративно-пускових апаратів. Для рухових ділянок кори характерний перш за все синтез збуджень різної модальності з біологічно значущими сигналами і мотиваційними впливами. Їм властива подальша, остаточна трансформація аферентних впливів в якісно нову форму діяльності, спрямовану на найшвидший вихід еферентних збуджень на периферію, тобто на апарати реалізації кінцевої стадії поведінки.
Передцентральна звивина є лише первинною (проекційною) зоною, виконавчим апаратом (вихідними воротами) мозкової кори. Природно, що склад рухових імпульсів, які надсилаються на периферію, має бути підготовленим, занесеним до певних програм, і тільки після такої підготовки рухова імпульсна програма може забезпечити потрібні доцільні рухи. Ця програма формується як у первинній моторній корі передцентральної звивини (поле 4), так і в корі надбудованих над нею вторинних (премоторні відділи, 5-те, 6-те, 7-ме і 8-ме поля) і третинних асоціативних зон (префронтальні відділи лобової кори, поля 9, 10, 11, 12, 46, 47).
Вторинні зони моторної кори морфологічно зберігають той же принцип “вертикальної організації”, що характерний для будь-якої моторної кори, але відрізняється незрівнянно більшим розвитком верхніх шарів кори, де переважають малі пірамідні клітини. Премоторна кора підлягає принципу спадної специфічності; у ній відсутня локальна соматотопічна проекція, а аксони пірамідних клітин цієї ділянки утворюють еференти, які перемикаються на великих підкоркових моторних утвореннях. Подразнення 5, 7, 8-го полів премоторної ділянки кори викликає не соматотопічно обмежені (локальні) здригання окремих м’язів, а цілі комплекси рухів, що мають системно організований характер (балістичні рухи очей у певну точку простору, повільні слідкуючі рухи очей, поворот голови, тулуба, направлені рухи кінцівок). Це вказує на те, що “командні” нейрони премоторної кори “організовують” окремі м’язові скорочення в цілісний руховий акт.
Третинні зони моторної кори являють собою блок програмування намірів, оцінки виконаних дій і корекції зроблених помилок, тобто апарат найскладніших форм регуляції цілісної поведінки. К. Прибрам (1975) вважає, що вони формують набір нейронних програм, що надають індивідуальному досвіду відому структуру, і будують “граматику” поведінки.
Переходячи на більш загальний рівень, можна сказати, що мозок підтримує організм, відчуваючи його потреби та спонукаючи до дій, необхідних для задоволення цих потреб. Однак людський мозок володіє також великими можливостями, які дозволяють порівнювати теперішнє з минулим. На основі багатьох сенсорних відомостей людина робить висновки, що майже в той же момент визначають, які рухові акти необхідні та які не потрібні. Численні паралельні опрацьовуючі системи дають людині велику кількість тлумачень поточної ситуації, які доповнюють можливу нестачу сенсорних даних за рахунок минулого досвіду.
С. Д. Максименко (2001) підкреслює: “Людська діяльність за сутністю є соціальною. Вона сформувалася історично, у процесі праці. Людина не лише пристосовується до умов життя, а й активно змінює їх відповідно до своїх потреб, що виникли й розвинулися історично… Особистість як суб’єкт діяльності, задовольняючи свої потреби, взаємодіє із середовищем, ставить перед собою певну мету, мотивує її, добирає засоби до її здійснення, виявляє фізичну і розумову активність у досягненні поставленої мети”.
Згідно з гіпотезою відомого американського нейрофізіолога Вернона Маунткасла (1981), в основі обробки сенсорної інформації та зворотної реакції на неї у всіх людей лежать схожі механізми. І все ж таки дія нейронних ансамблів кори півкуль великого мозку створює у кожної людини унікальне усвідомлення “Я”, відмінне від усіх інших.
Із усіх структур ЦНС кора півкуль великого мозку людини, імовірно, найскладніша за будовою та найбільш різноманітна за функціями. Однак безперечним та дивовижним залишається той факт, що певні ділянки кори посилають імпульси чітко визначеним частинам тіла або ж отримують імпульси від них. На дана карта чутливих та рухових ділянок кори, розроблена У. Пенфілдом та Т. Расмуссеном. На ній наглядно відображена важливість частин тіла людини. Надзвичайно велика частина кіркового кінця рухового аналізатора та аналізатора загальної чутливості віддана пальцям руки і особливо великому пальцю, а також ротові та органам мовлення (що в точності відповідає тим анатомо-функціональним особливостям, які виокремили людину з усього тваринного світу). Здатність людей зберігати та передавати інформацію, людська культура в цілому ніколи не могли б розвинутися без участі мови, а сучасна техніка і все, що створено людством, ніколи не з’явилось би на світ, якби у людини не було такої руки. У зв’язку з цим, мабуть, слід погодитися з Карлом Саганом (1986), який відмітив, що в певному розумінні карта чутливої частини кори великого мозку людини являє собою точний портрет усього людства.
Одним із сховищ центральних моторних програм у мозку є мозочок. У ньому зберігаються програми складних і автоматично виконуваних рухових актів, які сформувалися при житті людини. Було висловлене припущення, що зберігання інформації в мозочку здійснюється на модифікованих синапсах клітин Пуркіньє. В якості другої центральної структури, пов’язаної зі зберіганням моторних програм, дослідники розглядають базальні ядра, які мають відношення як до зберігання рухових програм вродженої поведінки, так і до рухових автоматизмів. Однак для того, щоб привести в дію рухові програми, необхідно, щоб у ці структури надійшов сигнал через систему ініціації рухів.
Однією із систем ініціації рухів є лімбічна система мозку, за допомогою якої здійснюється трансляція мотивації в дії, які пов’язані з втамуванням голоду, усуненням жаху і задовільненням інших потреб. Згідно з даними Г. Могенсона (1977), який розглядав механізми ініціації локомоцій, ця трансляція досягається за допомогою особливого шляху передачі сигналів від лімбічних структур до базальних ядер. При цьому важливою ланкою, яка зв’язує лімбічну систему з моторною, є розташоване в pars basalis telencephali прилегле ядро, n. accumbens. Його збудження сигналами з лімбічної системи, можливо, через механізми активації дофамінергічних шляхів, призводить до активації вроджених поведінкових програм.
Інша система ініціації руху через активацію моторних програм – це “когнітивний (або розумний) мозок”, представлений корою півкуль великого мозку. Він забезпечує ініціацію різних специфічних рухів у відповідності до інструкцій попереднього досвіду і навчання. Надходження сигналу від асоціативної кори до базальних ядер, які мають широкі сенсорні проекції, розгальмовує їх командні нейрони і тим самим активує моторні програми, головним чином рухові автоматизми, вивчені рухи. Ці програми через таламус також досягають моторної кори. Таким є шлях, через який “когнітивні програми” ініціюють програми руху, що зберігаються в ЦНС. За Г. Могенсоном, n. accumbens та n. caudatus формують сигнали відповідно від лімбічної системи (“емоційного мозку”) і від кори півкуль великого мозку (“когнітивного мозку”).
Таким чином, керування і контроль за рухом – досить складний процес. Він містить обробку інформації, яка надходить через прямі і зворотні зв’язки між різними відділами кори півкуль великого мозку, моторною корою, мозочком, базальними ядрами, а також стовбуром мозку та спинним мозком. Важлива роль належить пропріоцептивній та екстероцептивній аферентації. Рухова система організована за ієрархічним принципом з поступовим збільшенням складності сенсорної інтеграції. Рухова система працює як багаторівнева, при цьому кожний рівень структурно-функціональної організації мозку робить свій внесок таким чином, що діяльність більш високого рівня виникає не тільки сама по собі, але й як передумова появи та функціонування нового, вищерозташованого рівня.
Кровопостачання головного мозку
Кров надходить до головного мозку по двох парах артеріальних стовбурів: внутрішніх сонних артеріях (aa. carotis internus) і хребтових артеріях (aa. vertebrales).
Перш ніж потрапити в порожнину черепа, і хребтові і внутрішні сонні артерії утворюють S-подібні сифони, імовірно призначені для пом’якшення пульсових коливань. Внутрішні сонні артерії утворюють свої сифони в печеристих пазухах (печериста частина carotis interna). Хребтові артерії роблять це при виході з поперечних отворів атланта. S-подібні сифони у даному випадку утворюються за рахунок того, що ці артерії спочатку йдуть дозаду вздовж бічних поверхонь атланта, потім повертають догори і медіально і входять у порожнину черепа з обох боків довгастого мозку .
Хребтові артерії є головним джерелом кровопостачання стовбура мозку, мозочка та верхньошийних відділів спинного мозку. Вони являють собою перші гілки a. subclavia, які відходять зразу ж після виходу її з грудної порожнини (тобто на рівні перших ребер). На своєму шляху, у напрямку поперечних відростків сьомого шийного хребця, хребтові артерії розташовані позаду внутрішніх яремних вен, хребтових вен та нижніх шийних симпатичних вузлів.
Волокна цих вузлів супроводжують хребтові артерії, які піднімаються далі через поперечні отвори шістьох верхніх шийних хребців. Потім вони огинають бічні маси атланта і пронизують membrana atlantooccipitalis posterior. Артерії вступають у задню черепну ямку по обидва боки від довгастого мозку і конвергують спереду пірамід, утворюючи на рівні з’єднання довгастого мозку і моста єдиний стовбур основної артерії. Відповідно описаному вище ходу a. vertebralis в ній виділяють: передхребтову, поперечну, атлантову та внутрішньочерепну частини.
У порожнині черепа від внутрішньочерепної частини a. vertebralis відходять:
1) низхідні гілки, що утворюють передню спинномозкову артерію (a. spinalis anterior);
2) низхідні задні спинномозкові артерії (aa. Spinales posteriores);
3) артерії довгастого мозку – присередні мозкові гілки (rr. medullares mediales) та бічні мозкові гілки (rr. medullares laterales), які постачають відповідно присередні та бічні відділи кожної половини довгастого мозку (ці артерії досить варіабельні і можуть відходити також від основної артерії, або від задньої нижньої мозочкової артерії);
4) задні нижні мозочкові артерії (aa. inferiores posteriores cerebelli), перші гілки яких кровопостачають дорсолатеральні третини довгастого мозку.
Основна артерія (a. basilaris) є головним джерелом кровопостачання мосту, віддаючи до кожної його половини артерії мосту (aа. pontis), серед яких можна виділити короткі присередні гілки (rr. mediales), а також довші циркумферентні бічні гілки (rr. laterales). Присередні гілки ростральної частини основної артерії направляються до дна fossa interpeduncularis і кровопостачають не тільки найбільш верхні відділи pars basilaris pontis з обох боків від середньої лінії, але також присередні відділи верхньої половини tegmentum pontis, спускаючись через ростральні отвори substantia perforata posterior у міжніжковій ямці. Схожим чином, присередні гілки, що відходять від каудальної частини основної артерії в місці злиття хребтових артерій, огинають каудальні відділи pars basilaris pontis і проникають у сліпий отвір довгастого мозку (foramen caecum medullae oblongatae), тобто ростральний, що сліпо закінчується, кінець fissura mediana anterior між пірамідами довгастого мозку. Ці гілки постачають присередню частину каудальної половини tegmentum pontis. Бічні дві третини pars basilaris pontis et tegmentum pontis кровопостачаються rr. laterales (від aa. pontis).
Верхня мозочкова артерія (a. cerebelli superior) відходить від основної артерії безпосередньо перед розділенням останньої на дві задні мозкові артерії (а. cerebri posterior). Направляючись до мозочка, вона проходить до мосто-середньомозкового з’єднання через ростральну мосто-мозочкову щілину. Зробивши одну чи дві петлі, верхня мозочкова артерія виходить із щілини вздовж рострального краю мозочка.
Знаходячись у щілині, вона віддає гілки до верхньої ніжки мозочка. Інші дрібні гілки кровопостачають нижні горбки покрівлі середнього мозку і дорсолатеральну частину нижніх відділів покриву середнього мозку.
Крім верхньої мозочкової артерії, a. basilaris віддає також передню нижню мозочкову артерію (a. inferior anterior cerebelli), зона кровопостачання якої у порівнянні з верхньою і задньою нижньою мозочковими артеріями є незначною, обмежуючись корою і білою речовиною клаптика і суміжними вентральними листками мозочка.
Основним кровопостачальником переднього мозку є внутрішня сонна артерія, в якій (відповідно до її ходу) виділяють:
1) шийну частину (pars cervicalis);
2) кам’янисту частину (pars petrosa);
3) печеристу частину (pars cavernosa);
4) мозкову частину (pars cerebralis).
Увійшовши в підпавутинний простір, внутрішні сонні артерії (а саме їх мозкові частини) направляються дозаду під зорові нерви, а потім латерально до рівня зорового перехрестя, де вони розгортаються під прямим кутом, щоб увійти через бічну борозну у бічну ямку великого мозку. У місці повороту вони віддають задні сполучні артерії (aa. communicantes posteriores), які досягають проксимальних відділів задніх мозкових артерій (aa. cerebri posteriores), утворюючи разом з ними і переднім відрізком основної артерії задню дугу артеріального кола мозку, circulus arteriosus cerebri (кола Віллізія, circulus Willisii).
Внутрішні сонні артерії, перш ніж вони розділяються на передню та середню мозкові артерії, також віддають передні артерії судинного сплетення (aa. choroideae anteriores). Стовбури передніх мозкових артерій зразу ж направляються до середьої лінії і з’єднуються один з одним за допомогою передньої сполучної артерії (a. communicans anterior). Отже, цими артеріями замикається передня дуга артеріального кола мозку (кола Віллізія), яке має велике функціональне значення як пристосувальний механізм, що забезпечує безперебійне обхідне (колатеральне) кровопостачання головного мозку.
Описане нормальне коло Віллізія спостерігається менш ніж у половині випадків. У більшості людей воно дуже варіабельне, особливо його задня дуга (іноді може бути відсутня одна і навіть обидві задні сполучні артерії).
Передня мозкова артерія (a. cerebri anterior) йде вперед та присередньо, зближується з протилежною артерією і анастомозує з нею за допомогою передньої сполучної артерії. Цей анастомоз поділяє передню мозкову артерію на дві частини:
1) передсполучну частину (сегмент А 1), pars precommunicalis (segmentum A 1);
2) засполучну частину (сегмент А 2), pars postcommunicalis (segmentum A 2). мозолисто-крайова артерія (a. callosomarginalis)
Передсполучна частина передньої мозкової артерії віддає багато дрібних гілок, які входять у substantia perforata anterior і кровопостачають вентромедіальну половину передньої частини смугастого тіла, вентральні відділи передньої ніжки внутрішньої капсули, прозору перегородку та передню спайку. Одна з найпотужніших гілок до смугастого тіла зазвичай починається поблизу передньої сполучної артерії і йде у каудальному напрямку, щоб досягнути substantia perforata anterior. Ось чому Хюбнер описав її як поворотну артерію (a. recurrens). Інші дрібні гілки, які беруть початок біля a. communicans anterior, кровопостачають частину зорового перехрестя і прилягаючі ділянки зорових нервів.
Головними відгалуженнями засполучної частини a. cerebri anterior є: присередня лобово-основна артерія (a. frontobasalis medialis); лобова полюсна артерія (a. polaris frontalis); мозолисто-крайова артерія (a. callosomarginalis); навколомозолиста артерія (a. pericallosa). Гілки цих артерії кровопостачають кору лобової та тім’яної часток і верхніх ділянок потиличної частки. Навколомозолиста артерія є прямим продовженням передньої мозкової артерії і кровопостачає все мозолисте тіло, за винятком валика, який кровопостачає задня гілка мозолистого тіла (від сегмента Р4 задньої мозкової артерії).
Після відходження передньої мозкової артерії внутрішня сонна артерія перетворюється в середню мозкову артерію – найбільшу з усіх мозкових артерій.
Середня мозкова артерія (a. cerebri media) прямує вглиб бічної борозни мозку і кровопостачає кору острівцевої частки та більшу частину кори верхньобічної поверхні півкуль великого мозку. Стовбур середньої мозкової артерії поділяється на:
1) клиноподібну частину (сегмент М1), pars sphenoidalis (segmentum M1);
2) острівцеву частину (сегмент М2), pars insularis (segmentum M2).
Від початкової клиноподібної частини середньої мозкової артерії, розміщеної паралельно малим крилам клиноподібної кістки, відходять: передньобічні центральні артерії (aa. centrales anterolaterales); ближчі смугасті гілки (rr. proximales laterales striate); дальші бічні смугасті гілки (rr. distales laterales striate); гачкова артерія (a. uncalis); скронева полюсна артерія (a. polaris temporalis); передня скронева артерія (a. temporalis anterior).
Від острівцевої частини середньої мозкової артерії, розміщеної на поверхні острівцевої частки, відходять острівцеві артерії (aa. insulares), а також нижні та верхні кінцеві гілки. До нижніх кінцевих гілок (rr. termіnales inferiorеs) належать: передня, середня та задня скроневі гілки (r. temporalis anterior/medius/posterior); скронево-потилична гілка (r. temporooccipitalis); гілка кутової звивини (r. gyri angularis). До верхніх кінцевих гілок (rr. terminales superiores) належать: бічна лобово-основна артерія (a. frontobasalis lateralis); передлобова артерія (a. prefrontalis); артерія передцентральної борозни (a. sulci precentralis); артерія центральної борозни (a. sulci centralis); артерія зацентральної борозни (a. sulci postcentralis); передня та задня тім’яні артерії (a. parietalis anterior/posterior).
Задня сполучна артерія (a. communicans posterior) прямує до моста, де сполучається із задньою мозковою артерією (система підключичної артерії). Невеликі гілки задньої сполучної артерії живлять структури гіпоталамуса (сосочкове тіло, сірий горб, зорове перехрестя), передню третину таламуса, субталамус, частину задньої ніжки внутрішньої капсули, окоруховий нерв.
Задня мозкова артерія (a. cerebri posterior) є кінцевою гілкою основної артерії і складається з чотирьох частин:
1) передсполучної частини (сегмент Р1), pars precommunicalis (segmentum P1);
2) засполучної частини (сегмент Р2), pars post-communicalis (segment P2);
3) бічної потиличної артерії (сегмент Р3), occipitalis lateralis (segmentum P3);
4) присередньої потиличної артерії (сегмент Р4), occipitalis medialis (segmentum P4).
Передсполучна частина (сегмент Р1) розміщена до місця з’єднання задньої мозкової артерії та задньої сполучної артерії і віддає: задньоприсередні центральні артерії (aa. centrales posteromediales); короткі обвідні артерії (aa. circumferentiales breves); пронизну артерію таламуса (a. thalami perforans); горбкову або чотиригорбкову артерію (a. collicularis seu quadrigeminalis).
Жовтий колір – ділянка кровопостачання передньою мозковою артерією; червоний – середньою мозковою артерією; зелений – задньою мозковою артерією
Засполучна частина (сегмент Р2) задньої мозкової артерії огинає збоку ніжки мозку і виходить на дорсальну поверхню стовбура мозку. Ця частина віддає: задньобічні центральні артерії (aa. centrales posterolaterales); таламо-колінчасту артерію (a. thalamogeniculata); присередні та задні гілки судинного сплетення (rr. choroidei posteriores mediales et laterales); ніжкові гілки (rr. pedunculares).
До гілок бічної потиличної артерії (сегмент Р3) належать передні, проміжні та задні скроневі гілки (rr. temporales anteriores, intermedii et posteriors).
До гілок присередньої потиличної артерії (сегмент Р4) належать: задня гілка мозолистого тіла (r. corporis callosi dorsalis); тім’яна гілка (r. parietalis); тім’янопотилична гілка (r. parietooccipitalis); острогова гілка (r. calcarinus); потилично-скронева гілка (r. occipitotemporalis).
При раптовому порушенні кровопостачання мозку (тромбоз, емболія, розрив аневризми, травма) розвивається гостре порушення мозкового кровообігу (інсульт).
Кров з капілярів головного мозку надходить у поверхневі та глибокі вени великого мозку, які сполучаються з пазухами твердої оболони. Із пазух кров через внутрішні яремні, а потім через плечо-головні вени потрапляє у верхню порожнисту вену. Дуже невелика частина крові відтікає від головного мозку через венозні хребтові сплетення і через випускні вени (vv. emissariae), які з’єднують пазухи з венами губчатки (vv. diploicae) та венами зовнішніх покривів голови. Хід багатьох вен великого мозку незалежний від ходу артерій, які кровопостачають відповідні зони. Ці вени дренуються у верхню стрілову та поперечну пазухи. Інші вени йдуть майже паралельно відповідним артеріям і впадають у пазухи на основі черепа.
Поверхневі вени великого мозку (vv. superficiales cerebri) складаються з верхніх вен великого мозку (vv. superiores cerebri); середньої поверхневої вени великого мозку (v. media superficialis cerebri); нижніх вен великого мозку (vv. inferiores cerebri). Ці вени позбавлені клапанів, широко анастомозують між собою і через верхню та нижню сполучні вени (v. anastomotica superior et v. anastomotica inferior) сполучаються відповідно з верхньою стріловою та поперечною пазухами.
Глибокі вени великого мозку (vv. profundae cerebri) лежать у товщі мозку і збирають кров від його внутрішніх структур. До найбільш значних глибоких вен належать: основна вена (v. basilaris); велика вена великого мозку (v. magna cerebri) – вена Галена; внутрішні вени великого мозку (vv. internae cerebri).
Парна основна вена бере початок в ділянці substantia perforata anterior, огинає збоку ніжку мозку і вливається у велику вену великого мозку. Притоками основної вени є:
1) передні вени великого мозку (vv. anteriores cerebri), які супроводжують однойменні артерії;
2) середня глибока вена великого мозку (v. media profunda cerebri), в яку впадають острівцеві вени (vv. insulares);
3) нижні таламо-смугасті вени (vv. thalamostriatae inferiores);
4) вена нюхової звивини (v. gyri olfactorii);
5) нижня шлуночкова вена (v. ventricularis inferior);
6) нижня вена судинного сплетення (v. choroidea inferior), яка збирає кров від морського коника, зубчастої звивини та судинного сплетення нижнього рогу бічного шлуночка;
7) ніжкові вени (vv. pedunculares).
Внутрішні вени великого мозку формуються в ділянці міжшлуночкових отворів, йдуть назад через tela choroidea ІІІ шлуночка, зливаються одна з одною над lamina tecti середнього мозку і формують v. magna cerebri. До притоків vv. internae cerebri належать: верхня вена судинного сплетення (v. choroidea superior); присередня та бічна вени бічного шлуночка (v. medialis/lateralis ventriculi lateralis); бічні прямі вени (vv. directae laterals); задня вена прозорої перегородки (v. posterior septi pellucidi); верхня таламо-смугаста вена (v. thalamostriata superior). Верхня таламо-смугаста вена йде вперед у борозні між таламусом та хвостатим тілом, огинає передній горбок таламуса і в ділянці міжшлуночкового отвору вливається у v. interna cerebri. У верхню таламо-смугасту вену впадають вени хвостатого ядра (vv. nuclei caudati) та передня вена прозорої перегородки (v. anterior septi pellucidi).
Непарна v. magna cerebri має довжину до 1 см, залягає над lamina tecti середнього мозку і вливається у пряму пазуху. Крім vv. basilares та vv. internae cerebri вона приймає задню вену мозолистого тіла (v. posterior corporis callosi).
Зі сказаного зрозуміле те особливе значення, яке мають пазухи твердої мозкової оболони у забезпеченні венозного відтоку від головного мозку. Пазухи являють собою венозні, вистелені ендотелієм канали, у яких нема клапанів; залягають у товщі твердої оболони у місцях прикріплення її відростків до черепа і відрізняються від вен будовою своїх стінок. Непіддатливість стінок венозних пазух забезпечує вільний відтік венозної крові при зміні внутрішньочерепного тиску, що важливо для безперебійної діяльності головного мозку, чим і пояснюється наявність таких венозних пазух тільки у черепі.
Є такі пазухи твердої оболони головного мозку:
Верхня стрілова пазуха (sinus sagittalis superior) непарна, розміщена в основі falx cerebri вздовж усієї sulcus sinus sagittalis superioris від crista galli до protuberantia occipitalis interna і впадає у стік пазух; містить заглиблення – бічні затоки (lacunae laterales). Ця пазуха відкривається у стік пазух.
Стік пазух (confluens sinuum) – стік Герофіла – непарний, розміщений на protuberantia occipitalis interna, в місці злиття верхньої стрілової, прямої, поперечних та потиличної пазух.
Нижня стрілова пазуха (sinus sagittalis inferior) непарна, проходить дугоподібно спереду назад по нижньому вільному краю falx cerebri і впадає у пряму пазуху.
Пряма пазуха (sinus rectus) непарна, розміщена у місці прикріплення falx cerebri до tentorium cerebelli і впадає у стік пазух. Утворюється внаслідок злиття нижньої стрілової пазухи та великої вени великого мозку – вени Галена.
Потилична пазуха (sinus occipitalis) непарна, йде в основі falx cerebelli вздовж crista occipitalis interna.
Із стоку пазух венозна кров відтікає у праву і ліву поперечні пазухи (sinus transversae).
Поперечна пазуха (sinus transversae dexter et sinister) найбільша та найширша з усіх, розташована по задньому краю tentorium cerebelli в sulcus sinus transversi потиличної кістки, звідки спускається як сигмоподібна пазуха (sinus sigmoideus) в sulcus sinus sigmoidei і на рівні foramen jugulare переходить у внутрішню яремну вену (v. jugularis interna). Завдяки цьому поперечна та сигмоподібна пазухи служать головним колектором для всієї венозної крові від структур порожнини черепа. В них частково безпосередньо, частково опосередковано впадають всі інші пазухи.
Крайова пазуха (sinus marginalis) парна, йде по краю foramen magnum потиличної кістки, з’єднує потиличну пазуху ззаду з сигмоподібною пазухою збоку.
Печериста пазуха (sinus cavernosus) парна, розміщена по боках турецького сідла. Порожнина пазухи містить велику кількість перегородок, що надають їй печероподібного вигляду. Пазуха оточує внутрішню сонну артерію та відвідний нерв, а в її бічній стінці розміщені: окоруховий, блоковий, очний та верхньощелепний нерви.
Клино-тім’яна пазуха (sinus sphenoparietalis) парна, йде вздовж вільного краю малих крил клиноподібної кістки від внутрішньої поверхні тім’яної кістки до печеристої пазухи.
Передня та задня міжпечеристі пазухи (sinus intercavernosus anterior et sinus intercavernosus posterior) — пазухи Рідлея, що розміщені у турецькому сідлі і сполучають між собою печеристі пазухи попереду та позаду гіпофіза.
Верхня кам’яниста пазуха (sinus petrosus superior) парна, розміщена в однойменній борозні вздовж верхнього краю кам’янистої частини скроневої кістки, з’єднує печеристу пазуху із початковою ділянкою сигмоподібної пазухи .
Нижня кам’яниста пазуха (sinus petrosus inferior) парна, йде в однойменній борозні вздовж заднього краю кам’янистої частини скроневої кістки від печеристої пазухи до верхньої цибулини внутрішньої яремної вени (інколи впадає у сигмоподібну пазуху). На схилі основної частини потиличної кістки у вигляді сітки розміщене венозне основне сплетення (plexus basilaris), яке сполучає між собою печеристі, нижні кам’янисті, крайові пазухи, внутрішні хребтові венозні сплетення та хребтові вени.