Орган слуху та рівноваги
Вухо (auris) являє собою надзвичайно складний апарат, який сприймає звукові коливання, а також напрямок сили земного тяжіння та прискорення тіла людини, приймаючи тим самим участь у регулюванні довільних рухів. Вухо складається із зовнішнього, середнього та внутрішнього вуха і є периферійною частиною двох аналізаторів – слухового та статокінетичного.
Слід підкреслити, що кірковий кінець слухового аналізатора має надзвичайно важливе значення у розвитку пам’яті, інтелекту, мовної культури людини, у формуванні її мислення.
Зовнішнє вухо
Незважаючи на порівняно значні розміри, зовнішні структури вуха людини відіграють відносно невелику роль у процесах сприйняття звуку. За найпоширенішою точкою зору, функції зовнішнього вуха (аuris externa), яке включає вушну раковину (auricula), зовнішній слуховий хід (meatus acusticus externus) i барабанну перетинку (membrana tympani), обмежуються уловленням звукових хвиль. Зокрема, вушна раковина сприяє концентрації звуків, які приходять від певних ділянок простору в напрямку зовнішнього слухового ходу, бере участь в обмеженні потоку звукових сигналів, що надходять з тильної сторони голови, а також бере певну участь в ототопіці – визначенні напрямку звуків, які надходять з різних боків.
Вушна раковина (auricula) утворена складної форми еластичним вушним хрящем (cartilago auriculae), вкритим шкірою. Хрящ раковини прикріплюється до скроневої кістки за допомогою сполучнотканинних пучків, що утворюють три зв’язки вушної раковини (ligg. аuricularia). Передня вушна зв’язка (lig. аuriculare anterius) iде від arcus zygomaticus; верхня вушна зв’язка (lig. auriculare superius) iде від porus acusticus externus скроневої кістки; задня вушна зв’язка (lig. auriculare posterius) іде від processsus mastoideus скроневої кістки.
Вушна раковина, крім м’язів лиця (мімічних), що рухають її (m. temporoparietalis, m. auricularis anterior, m.auricularis superior, m.auricularis posterior), має ще слабкі рудиментарні нечисленні вольові м’язові волокна, які розміщуються пучками під шкірою на поверхнях вушної раковини у вигляді вушних м’язів (mm. auriculares).
У нижній третині вушної раковини, у так званій вушній часточці (lobulus auriculae), хрящового скелета нeмає, а складка шкіри півмісяцевої форми у цьому місці заповнена жировою тканиною. Вільний край вушної раковини, що завернутий всередину, називається завитком (helix). Передній кінець завитка, розміщений вгорі над зовнішнім слуховим ходом, називається ніжкою завитка (crus helices); нижній кінець завитка, розміщений внизу над вушною часточкою, називається хвостом завитка (cauda helices). Ніжка завитка має спрямовану вперед ость завитка (spina helices). Певний інтерес становить непостійний вушний горбок (tuberculum auriculae) – горбок Дарвіна, який іноді спостерігається у верхньозадній ділянці завитка. Інколи на місці вушного горбка є виступ завитка вушної раковини, спрямований назад, вгору та назовні – вушна верхівка (apex auriculae); вона відповідає верхівці вуха тварин.
Паралельно завитку на увігнутій ділянці вушної раковини проходить друге підвищення – протизавиток (antihelix). Протизавиток вгорі роздвоюється на дві ніжки протизавитка (crura antihelicis), які обмежовують неглибоку трикутну ямку (fossa triangularis). Завиток та протизавиток відокремлені один від одного борозною, що позначається як човен (scapha). Виступ вушної раковини – козелок (tragus) прикриває зовнішній слуховий хід спереду. Ззаду, на нижньому кінці протизавитка, якраз навпроти козелка, розташований схожий виступ – протикозелок (antitragus). Козелок та протикозелок розділені між собою вертикально розміщеною міжкозелковою вирізкою (incisura intertragica). Виразно виявлена заглибина вушної раковини, яка обмежована козелком, протикозелком і протизавитком, називається раковиною вуха(concha auriculae). Ніжка завитка розділяє раковину вуха на верхню меншу частину – човник раковини (cymba conchae) i нижню більшу частину – порожнину раковини (cavitas conchae); на дні останньої розташований зовнішній слуховий отвір.
Зовнішній слуховий хід (meatus acusticus externus) починається зовнішнім слуховим отвором (porus acusticus externus) i закінчується сліпо барабанною перетинкою, яка передає коливання повітря в зовнішньому вусі системі слухових кісточок середнього вуха. Зовнішній слуховий хід складається із зовнішньої слухової частини (яка становить третину його довжини і має назву хрящового зовнішнього слухового ходу (meatus acusticus externus cartilagines), і внутрішньої кісткової частини (яка розміщена всередині скроневої кістки). Основу хрящової частини ходу формує хрящ слухового ходу (cartilago meatus acusticis), який становить одне ціле з хрящем вушної раковини і має вигляд не замкненого вгорі жолоба, лійкоподібно звуженого у присередньому напрямі. Розміри зовнішнього слухового ходу зазнають індивідуальних коливань; середня довжина його у дорослого становить близько 25 мм. Просвіт зовнішнього слухового ходу має форму еліпса; його діаметр коливається від 6 до 9 мм. Meatus acusticus externus має складний S-подібний хід. Це треба мати на увазі при дослідженні його у живої людини: щоб випрямити кривизну хрящової частини зовнішнього слухового ходу, слід відтягнути вушну раковину дозаду і вгору.
Зовнішній слуховий хід разом з вушною раковиною можна порівняти з резонатором типу органної труби, закритої з одного боку. Власна частота його коливань залежить від довжини і форми комплексу “вушна раковина – зовнішній слуховий хід” і трохи відрізняється у різних людей. Резонансна частота міститься в діапазоні частот, які накладаються одна на одну, і рівна 3 кГц. На резонансних частотах акустичний тиск, який передається середньому та внутрішньому вухам, має максимальне значення. Посилення тиску на резонансній частоті зовнішнього вуха людини складає біля 10 дБ. Вважають, що існує зв’язок між мінімальним порогом чутності тонів визначеного діапазону і значенням резонансних частот зовнішнього вуха.
Слід зазначити також, що структури зовнішнього вуха відіграють певну захисну роль. Вони захищають барабанну перетинку від механічних і термічних впливів, забезпечують постійну температуру й вологість в ділянці барабанної перетинки. Шкіра початку слухового ходу має вушне волосся (tragi), яке у старих людей досягає особливого розвитку. Воно захищає вхід в meatus acusticus externus від зовнішніх впливів. Шкіра хрящового зовнішнього ходу має більшу товщину і містить багато сальних та сірчаних залоз. Вушна сірка (cerumen), що виділяється сірчаними залозами (видозміна потових залоз), є воскоподібною речовиною, що створює захисне покриття. У кістковій частині зовнішнього слухового ходу шкірний покрив, тісно зв’язаний з окістям, стає тоншим і на барабанну перетинку переходить тільки у вигляді епідермісу. Барабанна перетинка (membrana tympanica) відокремлює зовнішнє вухо від середнього вуха. Вона має площу 66–69,5 мм2, овальну форму і утворює з горизонтальною площиною кут у 45о, відкритий назовні, і майже такої самої величини кут утворює з серединною площиною; він відкритий назад (тому барабанна перетинка здається продовженням задньоверхньої стінки зовнішнього слухового ходу). Приблизно в центрі перетинка конусоподібно заглиблюється всередину барабанної порожнини, утворюючи пупок барабанної перетинки (umbo membranae tympanicae).
Барабанна перетинка побудована з тонкого шару волокнистої тканини, причому на периферії переважає колове спрямовання колагенових волокон, а в центрі – радіальне спрямовання. Зовні волокниста тканина перетинки вкрита епідермісом, а з боку барабанної порожнини – слизовою оболонкою. За допомогою волокнисто-хрящового кільця (anulus fibrocartilagineus) волокниста тканина на три чверті кола перетинки фіксована до барабанної борозни скроневої кістки. На одну чверть кола (саме вгорі, у вирізці pars tympanica) перетинка з’єднується з pars squamosa скроневої кістки. Тому в барабанній перетинці утворюється дві частини: 1) менша (завширшки близько 2 мм), що розміщена в ділянці згаданої вирізки і складає трикутної форми розслаблену частину (pars flaccida); 2) більша, з’єднана з барабанною борозною, утворює натягнуту частину (pars tensa). Слід підкреслити, що у pars flaccida барабанна перетинка не містить сполучнотканинних волокон.
Встановлено, що швидкість руху барабанної перетинки складає величина того ж порядку, що і швидкість зміщення часточок у плоскій хвилі повітря. При дуже слабких інтенсивностях звуку барабанна перетинка працює як нелінійна структура, генеруючи гармоніки частот, які її збуджують.
Середнє вухо
Середнє вухо (auris media) складається з барабанної порожнини, слухової труби та комірок соскоподібного відростка скроневої кістки. У барабанній порожнині містяться слухові кісточки (молоточок, коваделко, стремінце), м’язи, зв’язки, судини і нерви. Стінки її вистелені слизовою оболонкою, яка вкриває в більшій або меншій мірі названі органи, утворює певні складки і продовжується в слухову трубу та в комірки соскоподібного відростка скроневої кістки.
Барабанна порожнина (cavitas tympani) становить повітроносний простір неправильної форми, об’ємом близько 1 см3. Свого часу Фаллопій назвав її разом з барабанною перетинкою “tympanum” – “a similtudine cum tympano” (“за схожістю з барабаном”) і ця назва утрималась, хоча пізніше анатоми з’ясували, що такої схожості не існує. Барабанна порожнина розміщена всередині кам’янистої частини скроневої кістки. Для зручності опису в ній розрізняють 6 стінок.
Верхня – покривна стінка (paries tegmentalis) – утворена тонкою пластинкою компактної кісткової тканини, яка відповідає барабанному покриву на передній поверхні кам’янистої частини скроневої кістки та відокремлює барабанну порожнину від порожнини черепа. Вона містить надбарабанний закуток (recessus epitympanicus), який розташований над зовнішнім слуховим ходом. Окремо виділяють його верхню купольну частину (pars cupularis).
Нижня – яремна стінка (paries jugularis) – також тонка, обернена в бік яремної ямки, яка розташована на нижній поверхні кам’янистої частини скроневої кістки. Вона містить шилоподібний виступ (prominentia styloidea), який відповідає шилоподібному відростку скроневої кістки.
Передня – сонна стінка (paries caroticus) – відокремлює барабанну порожнину від сонного каналу і містить барабанний отвір слухової труби.
Задня – соскоподібна стінка (paries mastoideus) – містить вхід до печери (adіtus ad antrum). Соскоподібна печера (antrum mastoideum) сполучається з численними соскоподібними комірками (cellulae mastoideae), які розміщені у товщі соскоподібного відростка. На соскоподібній стінці розташовані зверху вниз декілька виступів: виступ бічного півколового каналу (prominentia canalis semicircularis lateralis); виступ каналу лицевого нерва (prominentia canalis facialis); пірамідальне підвищення (eminentia pyramidalis). Останнє розміщене на рівні вікна присінка, має порожнину всередині і маленький отвір на вершині, через який проходить сухожилок стремінцевого м’яза.
Біля входу до печери розташована ямка коваделка (fossa incudis), місце прикріплення задньої зв’язки коваделка. Між ямкою коваделка та пірамідальним підвищенням міститься задня пазуха (sinus posterior). Біля заднього краю барабанної перетинки на рівні пірамідального підвищення розташований барабанний отвір канальця барабанної струни (apertura tympanica canaliculi chordae tympani).
Присередня – лабіринтна стінка (paries labyrinthicus) – найскладніша і в той же час дуже важлива, бо відокремлює барабанну порожнину від внутрішнього вуха. Майже в центрі її виступає мис (promontorium) – підвищення, яке відповідає першому витку завитки. На поверхні мису можна бачити борозну мису (sulcus promontorii), в розгалуженнях якої залягають нерви барабанного сплетення. Над мисом є овальної форми вікно присінка (fenestra vestibuli), закрите основою стремінця. Нижче цього вікна розташована ямочка вікна присінка (fossula fenestrae vestibuli). Нижче ямочки міститься круглої форми вікно завитки (fenestra cochleae), закрите вторинною барабанною перетинкою (membrana tympanica secundaria). Вторинна барабанна перетинка відокремлює барабанну порожнину від барабанних сходів завитки та прикріплюється по краю вікна до гребеня вікна завитки (crista fenestrae cochleae). До вікна завитки веде ямочка вікна завитки (fossula fenestrae cochleae), позаду якої розташована підставка мису (subiculum promontorii), що обмежовує спереду глибоку ямку – барабанну пазуху (sinus tympani). Розташований над мисом на кінці перетинки canalis musculotubarius завиткоподібний відросток (processus cochleariformis) служить блоком, через який перекидається сухожилок м’яза-натягувача барабанної перетинки.
Бічна – перетинчаста стінка (paries membranaceus) – утворена барабанною перетинкою. По центру барабанної перетинки виразно виділяється пупок, до якого прилягає зрощена в цьому місці з перетинкою ручка молоточка. На зовнішній поверхні перетинки цій ручці відповідає молоточкова смуга (stria mallearis), яка тягнеться від пупка вгору до молоточкового виступа (prominentia mallearis), утвореного прилеглою до перетинки частиною бічного відростка молоточка. На внутрішній поверхні барабанної перетинки від основи ручки і головки молоточка до краю перетинки йдуть передня та задня молоточкові складки (plica mallearis anterior et plica mallearis posterior), які чітко обмежовують разом з краєм перетинки її розслаблену частину (pars flaccida).
До слухових кісточок (ossicula auditus) належать три найменші з усіх кісток скелета кісточки, які утворюють ланцюг, що з’єднує барабанну перетинку з вікном присінка на присередній стінці барабанної порожнини. Коливання барабанної перетинки призводять до руху першої в цьому ланцюгу кісточки – молоточка, потім коваделка, а потім кінцевого в ланцюгу стремінця. Основа стремінця, що укріплена у вікні присінка, у свою чергу призводить до руху перилімфи, яка заповнює сходи присінка та барабанні сходи у внутрішньому вусі. Звуковий тиск біля вікна завитки посилюється у 20 разів – це дуже важливо, оскільки рідина має значно більший акустичний опір, ніж повітря.
Слухові кісточки утримуються у барабанній порожнині зв’язками слухових кіcточок (ligg. ossiculorum auditus) і з’єднуються між собою за допомогою суглобів слухових кісточок (articulationes ossiculorum auditus), до яких належить коваделко-молоточковий суглоб (articulatio incudomallearis) та коваделко-стремінцевий суглоб (articulatio incudostapedalis). Стремінце прикріплюється до вікна присінка за допомогою барабанно-стремінцевого синдесмозу (syndesmosis tympanostapedalis).
Молоточок (malleus) має ручку молоточка (manubrium mallei), зовнішня поверхня якої від вільного кінця до бічного відростка молоточка прикріплюється до барабанної перетинки. Бічний відросток (processus lateralis) розташований на кінці ручки молоточка і відповідає молоточковому виступу барабанної перетинки. Масивна головка молоточка (caput mallei) повністю розміщена у верхній частині барабанної порожнини, де зчленовується з тілом коваделка. Між головкою та ручкою молоточка розміщена шийка молоточка (collum mallei). Витягнутий тонкий передній відросток (processus anterior) у новонароджених заходить у кам’янисто-барабанну щілину скроневої кістки, до якої прикріплюється передньою зв’язкою молоточка (lig. mallei anterius); у дорослих цей відросток редукується. Від головки молоточка до купольної частини надбарабанного закутка тягнеться верхня зв’язка молоточка (lig. mallei superius); шийку молоточка з барабанною борозною зв’язує бічна зв’язка молоточка (lig. mallei laterale).
Коваделко (incus) має тіло коваделка (corpus incudis), яке з’єднується з головкою молоточка за допомогою сідлоподібного суглоба, і дві ніжки – довгу та коротку. Довга ніжка (crus longus) прямує вертикально вниз позаду ручки молоточка і має на кінці сочевицеподібний відросток (processus lenticularis), за допомогою якого з’єднується зі стремінцем. Коротка ніжка (crus breve) спрямована дозаду і фіксується задньою зв’язкою коваделка (lig. incudis posterius) до розміщеної на задній стінці барабанної порожнини ямки коваделка. Верхня зв’язка коваделка (lig. incudis superius) прямує паралельно верхній зв’язці молоточка від тіла коваделка до купольної частини надбарабанного закутка.
Із трьох слухових кісточок стремінце (stapes) має найменші розміри; складається з головки, двох ніжок і основи. Головка стремінця (caput stapedis) має суглобову поверхню і з’єднується за допомогою art. incudostapedalis із сочевицеподібним відростком коваделка. Овальна основа стремінця (basis stapedis) прикріплюється до головки за допомогою передньої та задньої ніжок (crus anterius et crus posterius). Між ніжками та основою стремінця розташована тонка стремінцева перетинка (membrana stapedalis). Основа стремінця з’єднана з краями вікна присінка за допомогою кільцевої стремінцевої зв’язки (lig. anulare stapediale).
До розташованих в середньому вусі м’язів слухових кісточок (mm. ossiculorum auditus) належать: 1) м’язнатягувач барабанної перетинки (m. tensor tympani);
2) стремінцевий м’яз (m. stapedius).
М’яз-натягувач барабанної перетинки розміщений в однойменному півканалі скроневої кістки (semicanalis musculi tensoris tympani), який є верхньою частиною м’язово-трубного каналу. Довгий сухожилок цього м’яза виходить у барабанну порожнину, огинає розташований на її присередній стінці завиткоподібний відросток і прикріплюється до основи ручки молоточка.
Стремінцевий м’яз розміщений у порожнині пірамідального підвищення на задній стінці барабанної порожнини. Сухожилок м’яза виходить через маленький отвір на вершині пірамідального підвищення і прикріплюється до головки стремінця.
Традиційна точка зору на функції м’язів слухових кісточок полягає в тому, що їх рефлекторне скорочення, яке виникає при великій інтенсивності звуку, зменшує амплітуду коливань барабанної перетинки й слухових кісточок і таким чином зменшує коефіцієнт передачі рівня звукового тиску на внутрішнє вухо.
Між тим слід зазначити, що прихований період скорочення цих м’язів дуже великий (близько 10 мс), щоб захистити вухо від різких раптових звуків. Однак при тривалому перебуванні в умовах дії шумів скорочення м’язів слухових кісточок може мати принципове значення для акомодації звукопровідного апарату до звукових коливань різної амплітуди. Скорочення цих м’язів, особливо стремінцевого м’яза, відмічається також при орієнтовній реакції на появу нового подразника, при ковтанні та позіханні (коли змінюється тиск у барабанній порожнині), при жуванні та при мовленні людини. Це дозволяє розглядати активацію м’язів слухових кісточок не просто як захисний акустичний рефлекс, але й як важливу частину процесу продукування звуку, зворотного акустичного зв’язку й, відповідно, сприйняття біологічно важливих сигналів.
Слухова труба, tuba auditiva (auditoria) являє собою доволі довгий (в середньому 35 мм), вузький (завширшки 2 мм), трохи сплющений канал, який сполучає барабанну порожнину з носовою частиною глотки і служить для вирівнювання тиску повітря всередині барабанної порожнини по відношенню до зовнішнього атмосферного тиску, що необхідно для вільного вібрування барабанної перетинки. Цей орган дістав також назву труби Євстахія на честь відомого анатома XVI сторіччя Євстахія.
Слухова труба поділяється на меншу (одна третина труби), кісткову частину (pars ossea) та більшу (дві третини труби), хрящову частину (pars cartilaginea). Кісткова частина слухової труби розміщена у півканалі слухової труби (нижня частина canalis musculotubarius скроневої кістки). Стінку хрящової частини слухової труби формує хрящ слухової трубки (cartilago tubae auditivae), який складається з ширшої присередньої пластинки (lamina medialis) i вузької бічної пластинки (lamina lateralis). Загалом хрящ являє собою жолоб, відкритий донизу і вбік; дефект хряща доповнюється сполучнотканинною перетинчастою пластинкою (lamina membranacea). Найвужче місце слухової труби, розташоване в ділянці з’єднання хрящової частини з кістковою, має назву перешийка слухової труби (isthmus tubae auditivae). Звідси просвіт розширяється в обидва боки – як до барабанного отвору слухової труби (ostium tympanicum tubae auditivae), який відкривається на передній стінці барабанної порожнини, так і до глоткового отвору слухової труби (ostium pharyngeum tubae auditivae). Останній розміщений на боковій стінці носової частини глотки на висоті заднього кінця нижньої носової раковини, має форму лійки і обмежований двома валиками, задній з яких виявлений краще.
Внутрішнє вухо
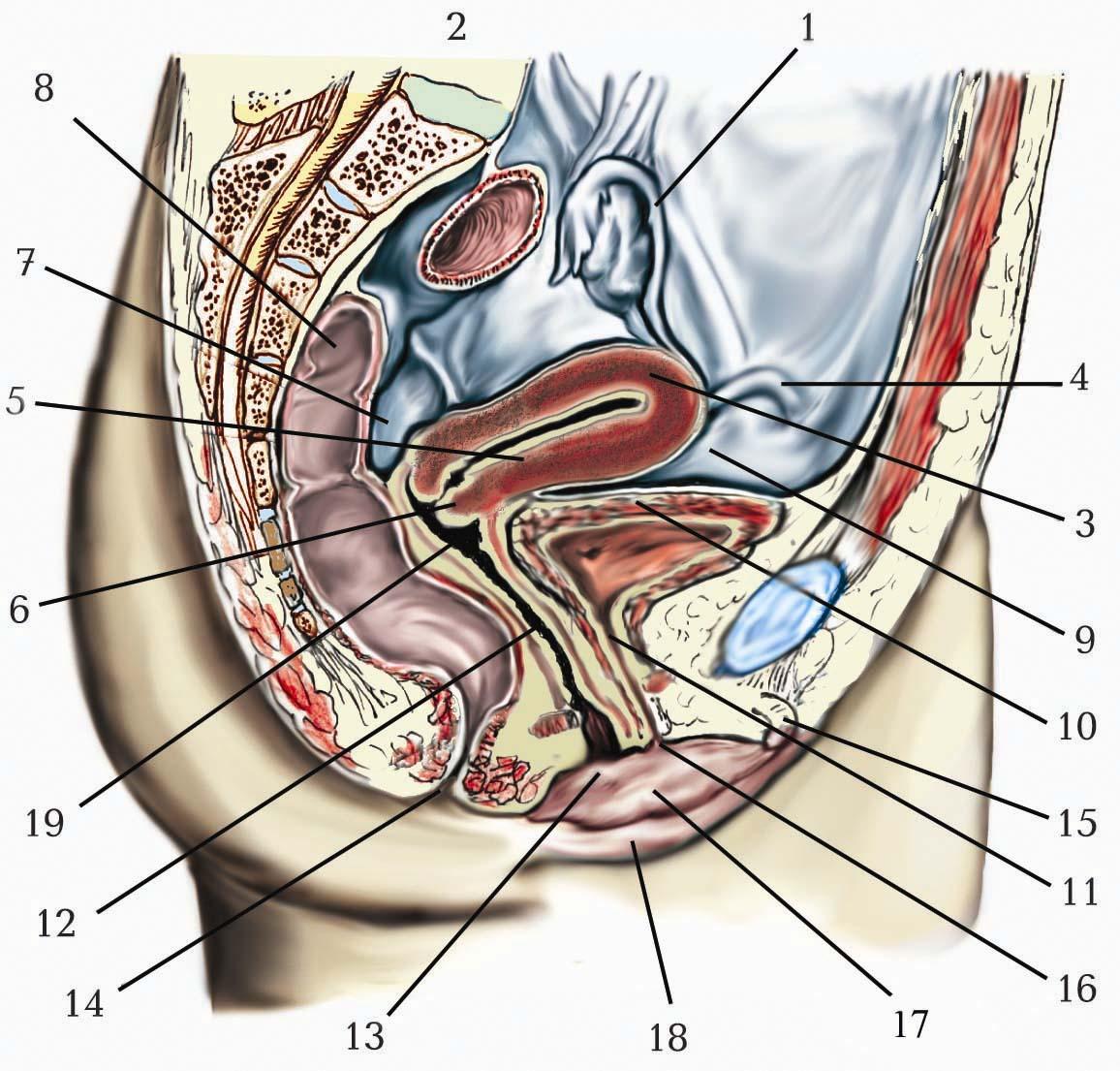
Внутрішнє вухо (auris interna) становить функціонально найважливіший і найскладніший за будовою відділ органа слуху та рівноваги. Воно складається з кісткового та перетинчастого лабіринтів, розміщених у кам’янистій частині (піраміді) скроневої кістки. Кістковий лабіринт розташований в губчастій кістковій речовині й містить всередині перетинчастий лабіринт, який повторює обриси кісткового лабіринту.
Кістковий лабіринт (labyrinthus osseus) представлений сукупністю різних за формою порожнин, стінка яких побудована зі щільної компактної кісткової речовини і вистелена всередині тонкою сполучнотканинною оболонкою. Кістковий лабіринт складається з трьох частин: центральне положення займає присінок, спереду від нього розташована завитка, ззаду містяться півколові канали.
Присінок (vestibulum) має вигляд порожнини неправильної форми, має виразний внутрішній рельєф і отвори на своїх стінках. На бічній стінці, оберненій до барабанної порожнини, розташоване овальної форми вікно присінка (fenestra vestibuli), закрите основою стремінця. Недалеко від нього, біля входу в завитку, міститься круглої форми вікно завитки (fenestra cоchleae), закрите вторинною барабанною перетинкою. На задній стінці присінка є 5 невеликих отворів півколових каналів. На передній стінці присінка розташований завитковий закуток (recessus cochlearis), в якому є один порівняно великий отвір входу в канал завитки. На присередній стінці присінка міститься еліптичний (маточковий) закуток, recessus ellipticus (utricularis), в якому є внутрішній отвір канальця присінка (apertura interna canaliculi vestіbuli). Невисокий гребінь присінка (crista vestibuli) відокремлює recessus ellipticus від розташованого нижче і більше наперед меншого та круглішого кулястого (мішечкового) закутка, recessus sphericus (saccularis). Крім того, присередня стінка містить решітчасті плями (maculae cribrosae), серед яких розрізняють:
1) верхню решітчасту пляму (macula cribrosa superior), через отвори якої проходять волокна маточковоампульного нерва (n. utriculoampullaris);
2) середню решітчасту пляму (macula cribrosa media), через отвори якої проходять волокна мішечкового нерва (n. saccularis);
3) нижню решітчасту пляму (macula cribrosa inferior), локалізовану в стінці задньої кісткової ампули; через її отвори проходять волокна заднього ампульного нерва (n. ampullaris posterior).
Півколові канали (canales semicirculares) мають дугоподібну форму і лежать у трьох взаємно перпендикулярних площинах:
1) горизонтальній – бічний півколовий канал (canalis semicircularis lateralis);
2) стріловій – передній півколовий канал (canalis semicircularis anterior), який проходить перпендикулярно до довгої осі піраміди;
3) лобовій – задній півколовий канал (canalis semicircularis posterior), який проходить паралельно задній поверхні піраміди.
Кожний канал має дві ніжки, з яких одна закінчується кістковою ампулою (ampulla ossea) і тому має назву ампульної кісткової ніжки (crus osseum ampullare). Інша вільна від ампули ніжка каналу називається простою кістковою ніжкою (crus osseum simplex). Ампульні кісткові ніжки відкриваються в присінок окремими отворами; сусідні прості кісткові ніжки переднього та заднього каналів зливаються разом і утворюють спільну кісткову ніжку (crus osseum commune), яка потім відкривається в присінок. Проста кісткова ніжка бічного півколового каналу відкривається в присінок самостійно. Саме тому три канали з шістьма ніжками з’єднуються з присінком за допомогою п’яти отворів.
Завитка (cochlea) має вигляд спірально закрученого каналу, який утворює два з половиною оберти навколо осі завитки, що йде майже горизонтально. Цей канал називається спіральним каналом завитки (canalis spiralis cochleae) і починається отвором на передній стінці присінка, а закінчується сліпо. Завитка має широку (7–9 мм) основу завитки (basis cochleae) і далі звужується у напрямку до купола завитки (cupula cochleae); загальна довжина спірального каналу становить біля 3 см. Осьовою частиною завитки є кісткове веретено завитки (modiolus cochleae), яке має конічну форму і оточене витками каналу. Всередині веретена проходять:
1) поздовжні канали веретена (canales longitudinales modioli) – містять у собі волокна завиткового нерва;
2) спіральний канал веретена (canalis spiralis modioli) – містить у собі спіральний вузол завитки.
Веретено завитки спірально обвивається кістковою спіральною пластинкою (lamina spiralis ossea), яка не досягає протилежної стінки каналу. Ця пластинка закінчується приблизно всередині поперечника спірального каналу завитки вільним краєм, від якого починається основна пластинка (lamina basilaris) та присінкова стінка (перетинка), paries (membrana) vestibularis, що досягають зовнішньої стінки каналу; за рахунок цього утворюється завиткова протока перетинчастого лабіринту. Кут між основною пластинкою та присінковою стінкою становить близько 45°, і вони формують відповідно нижню та верхню стінки завиткової протоки.
Кісткова спіральна пластинка разом із завитковою протокою обвивають веретено завитки, як гвинтові сходи свою вісь, і ділять просвіт спірального каналу на дві частини: верхню і нижню. Верхня, більша частина сполучається з присінком і має відповідну назву – сходи присінка (scala vestibuli). Нижня, менша частина через fenestra cochleae сполучається з барабанною порожниною, тому має назву барабанних сходів (scala tympani). Біля початку барабанних сходів, на дні їх, є маленький внутрішній отвір канальця завитки (apertura interna canaliculi cochleae). Зовнішній отвір цього канальця – apertura externa canaliculi cochleae – відкривається на нижній поверхні піраміди скроневої кістки.
Scala vestibuli et scala tympani є окремими ізольованими утвореннями; і тільки в кінці завитки вони сполучаються між собою через невеликий отвір завитки (helicotrema). Цей отвір обмежує гачок спіральної пластинки (hamulus laminae spiralis), який має вигляд загнутої гострої пластинки на кінці lamina spiralis ossea біля купола завитки.
Перетинчастий лабіринт (labyrinthus membranaceus) міститься всередині кісткового лабіринту, форму якого він частково копіює. Стінки перетинчастого лабіринту побудовані з волокнистої сполучної тканини і вистелені епітелієм. Через те що розміри перетинчастого лабіринту значно менші від розмірів кісткового лабіринту, між ними утворюється перилімфатичний простір (spatium perilymphaticum), заповнений прозорою лімфоподібною рідиною – перилімфою (perilympha). Перилімфа відтікає зі spatium perilymphaticum через водопровід завитки (aqueductus cochleae). Всередині перетинчастого лабіринту міститься подібна до перилімфи рідина, яка має назву ендолімфи (endolympha). Перетинчастий лабіринт є складною системою сполучених між собою порожнин і проток, серед яких виділяють присінковий лабіринт, півколові протоки та завиткову протоку .
Присінковий лабіринт (labyrinthus vestibularis) складається з маточки та мішечка. Маточка (utriculus) розміщена в recessus ellipticus (utricularis) кісткового лабіринту і є початком трьох півколових проток. Мішечок (sacculus) лежить в recessus sphericus (saccularis) кісткового лабіринту і сполучається з ductus cochlearis через сполучну протоку (ductus reuniens). Мішечок з’єднується з маточкою за допомогою маточково-мішечкової протоки (ductus utriculosaccularis), від якої починається ендолімфатична протока. Ендолімфатична протока (ductus endolymphaticus) проходить через водопровід присінка, досягає задньої поверхні кам’янистої частини скроневої кістки і закінчується сліпо ендолімфатичним мішечком (saccus endolymphaticus) у товщі твердої мозкової оболони.
На внутрішній стінці маточки та мішечка є білуватого кольору потовщення, які мають вид плям (maculae). Плями є рецепторами статокінетичного аналізатора, які сприймають силу земного тяжіння, статичне положення тіла людини та її прямолінійний поступальний рух без прискорення. Пляма маточки (macula utriculi) та пляма мішечка (macula sacculi) побудовані принципово однаково. Головним їх структурним компонентом є нейроепітелій, який складається з рецепторних та підтримуючих клітин. Рецепторні клітини мають волоски, занурені в желеподібну оболонку статичного піску (membrana statoconium), на поверхні якої розташовані кристалічні часточки карбонату кальцію, що мають назву статичного піску (statoconium). До рецепторних клітин підходять волокна відповідно n. utriculoampullaris et n. saccularis, які є гілками присінкового нерва (n. vestibularis).
Півколові протоки (ductus semicirculares) являють собою три трубки, які описують дугу в дві третини кола і розташовані у відповідних кісткових півколових каналах. В кожній кістковій ампулі півколових каналів містяться перетинчасті ампули: ampulla membranaceа anterior, ampulla membranacea posterior, ampulla membranacea lateralis. У кожній з перетинчастих ампул є по ампульному гребеню (crista ampullaris) .
Ампульний гребінь вкриває шар нейроепітеліальних рецепторних волоскових та підтримуючих клітин, вкритий, у свою чергу, желеподібним ампульним куполом (cupula ampullaris), в який занурюються волоски волоскових клітин. Знизу від ампульного гребеня проходить ампульна борозна (sulcus ampullaris), яка містить ампулярні гілки VIII пари черепних нервів, що йдуть до гребеня. Рецепторний апарат ампульних гребенів сприймає прискорені рухи тіла у різних напрямках.
Завиткова протока (ductus cochlearis) розміщена всередині спірального каналу завитки. Вона починається у присінку кісткового лабіринту присінковим сліпим кінцем (caecum vestibulare) і закінчується в ділянці купола завитки купольним сліпим кінцем (caecum cupulare). На поперечному розрізі завиткова протока має трикутний обрис і три стінки: зовнішню, нижню та верхню. Зовнішньою стінкою (paries externus) завиткова протока зростається зі стовщеним окістям спірального каналу. Це окістя вкрите спіральною зв’язкою (lig. spirale), яка має спіральний виступ (prominentia spiralis) і продовжується в основний гребінь нижньої стінки протоки. Спіральний виступ утворений сполучною тканиною і містить виступну судину (vas prominens). Вище спірального виступу у складі спіральної зв’язки розташована судинна смуга (stria vasculosa), яка вміщує внутрішньоепітеліальне капілярне сплетення, що продукує ендолімфу. Численними експериментальними дослідженнями доведено роль ушкодження судин stria vasculosa, які контролюють склад ендолімфи, в патогенезі нейросенсорної приглухуватості та глухоти. Вважають, що порушення іонного балансу ендолімфи призводить до змін рецепторних клітин спірального органа. Нижня стінка завиткової протоки має назву барабанної стінки (paries tympanicus) або спіральної перетинки (membrana spiralis). Більша частина нижньої стінки завиткової протоки утворена основною пластинкою (lamina basilaris). Основна пластинка тягнеться горизонтально від основного (спірального) гребеня, crista basilaris (spiralis), який утворений потовщенням окістя спірального каналу, до краю кісткової спіральної пластинки – спірального канату (limbus spiralis), а саме його нижнього довгого виступу – барабанної губи каната (labium limbi tympanicum). Основу lamina basilaris складають радіальні колагенові волокна різної довжини (0,04–0,5 мм), які порівнюють зі струнами, що під впливом коливань перилімфи тремтять залежно від своєї довжини в унісон з високими та низькими звуками. Верхня стінка піднімається вгору до зовнішньої стінки під кутом у 45° від кісткової спіральної пластинки і має назву присінкової стінки (перетинки), paries (membrana) vestibularis
На нижній стінці завиткової протоки розміщений спіральний орган (organum spirale). Цей орган ще називають кортієвим органом на честь італійського маркіза Альфонса Корті, який вперше описав його у 1854 році, коли працював на кафедрі анатомії під керівництвом відомого австрійського анатома І. Гіртля. Із внутрішнього та зовнішнього боків спіральний орган обмежований внутрішньою та зовнішньою спіральними борознами (sulcus spiralis internus et sulcus spiralis externus).
Спіральний орган побудований з потрійного родуклітин:
1) внутрішніх та зовнішніх волоскових клітин;
2) внутрішніх та зовнішніх фалангових клітин; 3) підтримуючих клітин.
Клітини спірального органа вкриті сітчастою перетинкою (membrana reticularis), яка сформована маленькими відростками (фаланговими відростками) фалангових клітин. У перфорації сітчастої перетинки заходять волоски волоскових рецепторних клітин. Над спіральним органом розташована покривна мембрана (membrana tectoria), яка прикріплена до верхнього коротшого виступу спірального каната – присінкової губи каната (labium limbi vestibulare). У місці прикріплення покривної пластинки на поверхні присінкової губи каната виступають гребенеподібні ряди клітин, які мають назву слухових зубців (dentes acustici). В основній пластинці є отвори, які містять нерви, що йдуть від волоскових клітин до спірального вузла завитки. До цього часу остаточно не з’ясовано, чи доторкаються волоски волоскових клітин до покривної мембрани, чи просто зміщуються під часколивань ендолімфи, які викликані коливаннями перилімфи; але не викликає сумнівів, що чутливі волоскові клітини спірального органа трансформують механічну енергію звукових коливань в енергію нервового імпульсу.
Звукопровідний та звукосприймаючий апарати вуха. Теорії слуху
До звукопровідного апарату вуха належать структури зовнішнього та середнього вуха, а також перилімфа та ендолімфа внутрішнього вуха. Основне призначення зовнішнього вуха полягає у проведенні звукових хвиль до барабанної перетинки. У свою чергу, основна роль барабанної перетинки та системи слухових кісточок полягає в трансформації звукових коливань великої амплітуди та малої сили в коливання рідин внутрішнього вуха з малою амплітудою та великою силою.
Кінцевим етапом проведення звуку до внутрішнього вуха є рух основи стремінця у вікні присінка, який передається перилімфі scala vestibuli. Коливання перилімфи scala vestibuli досягають купола завитки, де вони через отвір завитки передаються перилімфі scala tympani. По перилімфі scala tympani звукові коливання повертаються до присінка і гасяться вторинною барабанною перетинкою, яка закриває fenestra cochleae. Від перилімфи звукові коливання передаються ендолімфі завиткової протоки і сприймаються волосковими рецепторними клітинами спірального органа, причому велике значення в цьому процесі має основна пластинка.
Основна пластинка (lamina basilaris) має довжину 35 мм, а ширина її (і довжина колагенових волокон) збільшується у напрямку від вікна присінка до купола завитки. Така тонотопічна організація основної пластинки дає змогу людині сприймати коливання повітря у діапазоні від 16 до 20000 Гц. Вухо людини нечутливе до коливань частотою менше 16 Гц (інфразвукові коливання), тому звуки, що утворюються рухами більшості його власних органів (рухи м’язів, нутрощів, биття серця і таке інше) залишаються нечутними.
За традицією, яка склалася упродовж багатовікової історії вивчення слухових функцій людини і тварин, основні інтереси дослідників концентрувались навколо проблеми саме слухового аналізу частоти звукових коливань. У 1863 році Г. Гельмгольц виступив з резонансною теорією слуху, яка базувалась на думці, що окремі волокна основної пластинки настроєні, як струни, на різні звукові частоти.
У подальшому було встановлено, що при дії хвилі звуку, яка передається до fenestra vestibuli за допомогою слухових кісточок, на основній пластинці виникає так звана бігуча хвиля. Вибухання пластинки переміщується від cupula cochleae до fenestra cochleae. Відстань, яку проходить бігуча хвиля, і швидкість, з якою вона розповсюджується, визначається частотою звукової хвилі. Оскільки основна пластинка розширюється від fenestra cochleae до cupula cochleae, а жорсткість її збільшується у цьому напрямку майже в 100 разів, то амплітуда бігучої хвилі зменшується по мірі віддалення від cupula cochleae. Місце максимального відхилення основної пластинки пов’язане із частотою звуку: для звуків високої частоти воно розташоване біля fenestra cochleae, для звуків низької частоти – біля cupula cochleae. Таким чином, різні звукові частоти, які перетворюються в амплітуди відхилення пластинки, локалізовані в різних її місцях. Це перетворення частоти коливань у місце максимального відхилення пластинки лежить в основі одного із найбільш розповсюджених варіантів резонансної теорії – теорії “місця” .
Саме по собі місце максимального коливання основної пластинки, яке визначається механічними і гідродинамічними властивостями структур внутрішнього вуха, природно, не є достатнім для сприйняття висоти тону. Необхідно, щоб існували спеціальні нервові механізми (звукосприймаючий апарат), що трансформують механічний процес у просторовий малюнок активності слухових центрів.
В межах теорії періодичності сприйняття висот В. Резерфорд у 1886 році висунув “телефонну” теорію слуху. У відповідності з нею, частота звукової хвилі передається в мозкові центри, як звукові (електричні) коливання в телефонному проводі. Вразливим положенням цієї теорії є те, що поодинокі нервові волокна можуть розряджатися тільки до визначених граничних частот, а саме – до 800–1200 Гц. Верхня границя слуху людини і більшості ссавців лежить у діапазонах частот понад 10 000 Гц (а у дрібних гризунів та кажанів – до 80–150 кГц).
У 1948 році “телефонна” теорія була замінена теорією “залпів”, запропонованою Е. Уівером та С. Бреєм, першовідкривачами так званого “мікрофонного ефекту завитки” (1930). Вони висловили думку, що для визначення висоти тону важлива не частота розряду в поодинокому нервовому волокні, а сумарна частота розряду в ансамблі нейронів. “Залп” ансамблів нейронів може відтворювати частоти звуку, значно вищі, ніж ті, що доступні одиничному нервовому волокну. Новіші дослідження, виконані групою американських учених на чолі з Н. Кіакаганом (1960) на одиничних нервових волокнах із застосуванням обчислювальної техніки, довели, що все-таки в розряді одиничного нервового волокна, а саме в його міжімпульсних інтервалах, можуть знаходити відображення частоти до 5000 Гц.
Завершуючи огляд теорій слуху, слід зазначити, що нині прийнято говорити про дуалізм механізмів сприйняття висоти тону: в ділянці високих частот найбільше підходить “принцип місця”, в ділянці низьких частот – модифікований “принцип залпів”.
Провідний шлях аналізатора слуху
Тіло першого нейрона шляху слухового аналізатора лежить у завитковому вузлі, або спіральному вузлі завитки. Завитковий вузол розташований у спіральному каналі веретена. Периферійні відростки (дендрити) біполярних нейронів, які утворюють завитковий вузол, через отвори в базальній пластинці зв’язані з рецепторними клітинами спірального органа. Центральні відростки (аксони) нейронів завиткового вузла утворюють завитковий нерв (n. cochlearis), який приєднується до присінкового нерва (n. vestibularis), на своєму шляху через meatus acusticus internus до мосто-мозочкового кута. В цьому місці обидва нерви, які утворюють присінково-завитковий нерв (n. vestibulocochlearis) (VIII) вступають у стовбур мозку безпосередньо позаду pedunculus cerebellaris inferior.
Волокна n. cochlearis входять у стовбур мозку на рівні переднього завиткового ядра VIII пари черепних нервів (n. cochlearis anterior) та розподіляються на висхідні та низхідні волокна. Висхідні волокна йдуть до заднього завиткового ядра VIII пари черепних нервів (n. cochlearis posterior), а низхідні гілки – до n. cochlearis anterior. Завитка досить точно проектується в завитковий ядерний комплекс: волокна, які передають нервові імпульси з базальних витків, закінчуються в дорсоприсередній частині ядер, а волокна, які передають нервові імпульси з верхніх витків завитки, закінчуються в центроприсередній частині; такий впорядкований розподіл аферентних волокон створює основу організації завиткових ядер, у відповідності до тональних частот.
Аксони нейронів n. cochlearis anterior переходять напротилежну сторону у вигляді широкої пластинки волокон та утворюють разом з розташованими між ними нейронами трапецієподібне тіло (corpus trapezoideum). Волокна, які беруть початок від n. cochlearis posterior, переходять на протилежну сторону частково у складі мозкових смуг четвертого шлуночка (striae medullares ventriculi quarti), частково у складі ретикулярної формації. Волокна від завиткових ядер (тіло ІІ нейрона слухового шляху) утворюють бічну петлю (lemniscus lateralis). Багато волокон lemniscus lateralis безпосередньо йдуть до нижніх горбків пластинки покрівлі (чотиригорбкової пластинки) середнього мозку. Деякі волокна lemniscus lateralis утворюють синапси в проміжних ядрах слухового шляху (тіла ІІІ нейрона слухового шляху):
1) у верхньому оливному ядрі (дорсальне ядро трапецієподібного тіла, BNA) n. olivaris superior (n. dorsalis corporis trapezoidei, BNA);
2) у передньому ядрі трапецієподібного тіла (вентральне ядро трапецієподібного тіла, BNA), n.anterior corporis trapezoidei (n. ventralis corporis trapezoidei, BNA);
3) у присередньому верхньому оливному ядрі (n. olivaris superior medialis);
4) в ядрах бічної петлі (nuclei lemnisci lateralis). Ядра бічної петлі (заднє, проміжне, переднє) представлені групами нейронів, які розподілені по ходу lemniscus lateralis.
Верхнє оливне ядро характеризується вираженою тонотопічною організацією. Присереднє верхнє ядро (оливне) отримує волокна від завиткових ядер обох сторін і є вставним ядром у системі слухових волокон, яке сприяє направленому слуху. З’єднання волокон n. cochlearis posterior VIII пари черепних нервів з ядром nucleus nervi abducentis забезпечує рефлекторні рухи очима у відповідь на звуковий подразник.
Виходячи зі сказаного вище, lemniscus lateralis є достатньо складним утворенням, що сформоване аксонами ІІ та ІІІ нейронів, які йдуть гомота гетеролатерально, тому ушкодження однієї lemniscus lateralis не викликає односторонньої глухоти. У цьому випадку звичайно має місце невелике зниження слуху з протилежного боку (гіпоакузія) та деяке порушення впізнання напрямку звуку.
Більшість волокон lemniscus lateralis закінчуються в ядрах нижнього горбка пластинки покрівлі середнього мозку. Останні є релейною станцією для слухових рефлексів, від них ідуть висхідні волокна до верхніх горбків та низхідні волокна до спинного мозку (tractus tectospinalis), до стовбура мозку (tractus tectobulbaris) та до мозочка (fasciculus tectocerebellaris). Tractus tectospinalis йде до мотонейронів передніх рогів спинного мозку та забезпечує поворот голови у бік джерела звуку або від нього. Tractus tectobulbaris іде до рухових ядер різних черепних нервів, у тому числі – до рухового ядра лицевого нерва (для регуляції тонуса стремінцевого м’яза). Ядра нижніх горбків віддають також колатералі до висхідної активуючої системи ретикулярної формації та сприяють організації процесу прокидання від сну.
Через ручку нижнього горбка (brachium colliculi inferioris) волокна слухового шляху досягають ядер присереднього колінчастого тіла (nuclei corporis geniculati medialis) (тіла IV нейрона слухового шляху). Нещодавні дослідження показали, що corpus geniculatum mediale отримує також соматосенсорні волокна зі спинного мозку та волокна від мозочка. Таким чином, присереднє колінчасте тіло – це важлива для слухової системи синтетична ділянка, яка має зв’язки з іншими функціональними системами. Слід особливо підкреслити, що більшість волокон слухового шляху беруть початок від оливних ядер моста та ядер розташованого в мосту трапецієподібного тіла і досягають corpus geniculatum mediale без попереднього синапсування в нижньому горбку пластинки покрівлі середнього мозку.
Від corpus geniculatum mediale волокна слухового шляху йдуть у складі слухової променистості (колінцево-скроневих волокон), radiatio acustica (fibrae geniculotemporale), через підсочевицеподібну частину (pars sublentiformis) задньої ніжки внутрішньої капсули до кори, розташованих на внутрішній поверхні верхньої скроневої звивини поперечних скроневих звивин – gyrus temporalis transversus anterior et gyrus temporalis transversus posterior. У цих звивинах (цитоархітектонічне поле 41 за Бродманом) розташовується первинне слухове поле, або кірковий кінець слухового аналізатора (за І. П. Павловим). Первинне слухове поле кори великого мозку людини має тонотопічну організацію, причому завитка в розгорнутому вигляді спроектована на поперечні скроневі звивини зверху вниз відповідно від basis cochleae до cupula cochleae. Прилеглі асоціативні слухові поля також мають тонотопічну організацію, представлену на.
Первинне слухове поле 41 оточене частково полем 42, збоку від якого знаходиться поле 22, яке займає зов-нішню поверхню верхньої скроневої звивини. У цих вторинних ділянках слухові стимули аналізуються, ідентифікуються та порівнюються з первинними в слуховій пам’яті. Вони також інтерпретуються та впізнаються в якості шумів, тонів, мелодій, голосних та приголосних звуків, слів та речень – тобто символів мови. У випадку ушкодження вказаних кіркових полів у домінантній півкулі великого мозку втрачається властивість впізнавати звуки та розуміти власну мову (сенсорна афазія).
Кіркова частина слухового аналізатора грає першоступеневу роль у слуховій пам’яті (в тому числі у пам’яті слухових емоцій), слухо-мовних взаємовідносинах. Слуховий аналізатор є аферентною частиною, ланкою в надзвичайно складній рефлекторній дузі мовлення. Акт мовлення включає декілька етапів становлення мови в індивідуальному житті: сприйняття мовних звуків у ранньому дитячому віці, накопичення елементарних звуко-мовних образів, потім пізнання мовно-предметних співвідношень, сприйняття змісту мовних сигналів тощо. Процес пізнання іде в комплексі отриманих зорових, тактильних та інших подразнень, переходить нарешті від сенсорної до рухової функції органів мовлення.
Провідний шлях статокінетичного аналізатора
Три аналізатори забезпечують збереження рівноваги та підтримання певного положення тіла людини у просторі:
1) статокінетичний аналізатор;
2) пропріоцептивний аналізатор;
3) зоровий аналізатор.
Що стосується статокінетичного аналізатора, то слід зазначити, що адекватним подразником рецепторних утворень, закладених у перетинчастих ампулах півколових проток, є кутові прискорення, які спричиняють інерційне зміщення ендолімфи, деформацію рухомих частин ампульних гребенів та збудження волоскових клітин під час обертання голови. Адекватним подразником рецепторних утворень, закладених у маточці та мішечку присінка, є сили гравітації, що чинять постійний тиск на статичний пісок і його желеподібну оболонку, а також прямолінійні прискорення та відцентрові сили, що викликають тангенціальні зміщення желеподібної оболонки статичного піску та збудження волоскових нейроепітеліальних клітин.
Тіло першого нейрона шляху статокінетичного аналізатора розміщене у присінковому вузлі (ganglion vestibulare), який розміщений на дні внутрішнього слухового ходу. Дендрити біполярних нейронів присінкового вузла утворюють його верхню (pars superior) та нижню (pars inferior) частини. Pars superior продовжується у маточково-ампульний нерв (n. utriculoampullaris), який розділяється на маточковий нерв (n.utricularis) (починається від рецепторів плями маточки) та бічний ампульний нерв (n. ampullaris lateralis) (починається від рецепторів ампульних гребенів передньої та бічної півколових проток). Pars inferior продовжується у мішечковий нерв (n. saccularis) (починається від рецепторів плями мішечка) та задній ампульний нерв (n. ampullaris posterior) (починається від рецепторів ампульного гребеня задньої півколової протоки).
Аксони біполярних нейронів присінкового вузла утворюють присінковий нерв (n. vestibularis), який приєднується до завиткового нерва (n. cochlearis) і разом з ним утворює присінково-завитковий нерв (n. vestibulocochlearis), який виходить з піраміди скроневої кістки через внутрішній слуховий отвір і вступає в мозкову речовину моста у мосто-мозочковому куті.
Нервові імпульси від рецепторних клітин різних частин перетинчастого лабіринту йдуть по нервових волокнах до різних ділянок присінкового ядерного комплексу. Нервові волокна, що передають імпульси від macula sacculi, закінчуються в бічній частині nucleus vestibularis inferior, а від macula utriculi – в присередній частині nucleus vestibularis inferior та в бічній частині nucleus vestibularis medialis. Нервові волокна, що передають імпульси від ампульних гребенів півколових проток, закінчуються головним чином у nucleus vestibularis superior та у верхній частині nucleus vestibularis medialis.
Певні групи нейронів присінкового ядерного комплексу реагують на прямолінійне прискорення, а інші – на доцентрове прискорення; деякі нейрони реагують на обертання у гомолатеральний бік, а інші – на обертання у гетеролатеральний бік. Лівий та правий присінкові ядерні комплекси пов’язані між собою комісуральними волокнами, тому певна група їх нейронів реагує на подразнення гетеролатеральних перетинчастих лабіринтів.
Крім волокон безпосередньо від присінкового вузла, archicerebellum отримує волокна від верхнього, присереднього та нижнього присінкових ядер. Archicerebellum посилає еферентні імпульси назад у комплекс присінкових ядер, а також у спинний мозок до його мотонейронів безпосередньо по вершинно-спинномозковому шляху (який іде у бічному канатику спинного мозку), та опосередковано через мозочково-ретикулярні та ретикуло-спинномозкові зв’язки.
Від бічного присінкового ядра починається важливий екстрапірамідний шлях – бічний присінково-спинномозковий шлях (tractus vestibulospinalis lateralis). Він спускається гомолатерально в передньому канатику до альфата гамма-мотонейронів спинного мозку, досягаючи крижових сегментів. Цей шлях здійснює координацію рухів м’язів при розгинанні у суглобах і забезпечує тонус м’язів-розгиначів, достатній для підтримки рівноваги.
Від присереднього присінкового ядра починається ще один екстрапірамідний шлях – присередній присінково-спинномозковий шлях (tractus vestibulospinalis medialis), який у стoвбурі мозку з кожного боку приєднується до присереднього поздовжнього пучка. Цей шлях спускається в передньому канaтику спинного мозку, забезпечує зв’язок присінкових ядер з мотонейронами шийних сегментів і закінчується у грудному відділі спинного мозку у вигляді пучка крайової борозни (fasciculus sulcomarginalis). Присередній присінково-спинномозковий шлях впливає на тонус м’язів шиї у відповідності з різними положеннями голови. Можливо також, що він бере участь у забезпеченні рефлексів, які сприяють підтриманню рівноваги шляхом здійснення початкових компенсаторних рухів верхніми кінцівками.
Статокінетична система повинна мати зв’язки з корою півкуль великого мозку, оскільки людина усвідомлює своє положення та його зміни у просторі. Однак присінково-кіркові провідні шляхи повністю ще не ідентифіковані. Вважається, що від присінкових ядер прямує перехрещений шлях до таламуса. В останньому розміщені тіла ІІІ нейронів шляху статокінетичного аналізатора, аксони яких досягають кори верхньо-бічної поверхні скроневої частки (головним чином середньої та нижньої скроневих звивин). Дифузний характер кіркового кінця статокінетичного аналізатора пов’язують з тим, що положення людини у просторі може бути оцінене нею точно тільки за умови негайної реєстрації зорових, пропріоцептивних та вестибулярних сигналів у ЦНС та їх інтеграції один з одним.
Розвиток органа слуху та рівноваги
Вухо як анатомічне утворення складається з органа слуху та рівноваги, які розвиваються із зачатків різного походження.
Внутрішнє вухо людини розвивається з ектодерми головного кінця зародка на рівні заднього мозку. Першим закладається перетинчастий лабіринт у вигляді слухового пухиря.
У зародка людини на початку третього тижня розвитку по боках ще відкритої нервової пластинки виникають незначні потовщення головної ектодерми – плакоди. В середині третього тижня розвитку слухові плакоди вже добре виражені, а в кінці третього тижня кожна слухова плакода набуває форми чітко окресленого потовщення ектодерми по боках заднього мозку, що розвивається. Упродовж четвертого тижня розвитку плакода інвагінує, і утворюється слухова ямка. Слухова ямка поступово заглиблюється, потім її краї замикаються, вона відділяється від ектодерми та перетворюється у заповнений ендолімфою слуховий пухирець.
У місці відокремлення слухового пухирця від ектодерми формується трубчастий відросток пухирця – ендолімфатична протока. Дорсальна частина слухового пухирця, яка зв’язана з ендолімфатичною протокою, є закладкою присінкового лабіринту, а тонша вентральна частина – закладкою завиткової протоки.
В кінці шостого тижня розвитку з присінкової частини слухового пухирця починають розвиватися три півколові протоки. Одночасно з утворенням півколових проток присінкова частина слухового пухирця поділяється на більш дорсальну ділянку, яка називається маточкою, та більш вентральну ділянку – мішечок.
Упродовж перших шести тижнів розвитку зародка людини частина перетинчастого лабіринту, з якого розвивається завиткова протока, швидко подовжується. Ріст цієї частини лабіринту продовжується з наростаючою швидкістю упродовж 7-го та 8-го тижнів, в результаті чого утворюється спіраль з 2,5 обертами. По мірі формування завиткової протоки її первісне широке з’єднання з присінковим лабіринтом перетворюється в тонку сполучну протоку (ductus reuniens).
Вже на ранніх стадіях свого розвитку слуховий пухирець індукує оточуючу мезенхіму мезодермального походження до формування слухової капсули. Слухова капсула проходить сполучнотканинну стадію розвитку та перетворюється в хрящ, який повторює контури перетинчастого лабіринту, що розвивається, і називається хрящовим лабіринтом. Між стінками перетинчастого та хрящового лабіринтів утворюється перилімфатичний простір, заповнений рідиною (перилімфою), аналогічною спинномозковій рідині. На пізніших стадіях пренатального онтогенезу хрящова тканина замінюється кістковою і утворюється кістковий лабіринт.
На ранніх стадіях ембріогенезу ентодерма первинної глотки утворює парні бічні випинання – зяброві кишені, а ектодерма утворює відповідні їм присередні випинання – зяброві щілини; при цьому розташована між ними мезенхіма мезодермального походження сегментується на зяброві дуги. Перша зяброва кишеня простягається латерально до тих пір, поки вистеляюча його ентодерма не прийде у контакт з ектодермою дна першої зябрової щілини, утворюючи зяброву пластинку. Із кінцевої частини І зябрової кишені виникає барабанна порожнина, а проксимальна частина в майбутньому звужується та утворює слухову трубу. Крім цього, ентодерма І зябрової кишені вростає в закладку скроневої кістки та утворює соскоподібну печеру і соскоподібні комірки.
І зяброва щілина дає початок розвитку порожнини вушної раковини та зовнішнього слухового ходу. Сліпий кінець І зябрової кишені (закладка барабанної порожнини) відходить від поверхні й оточується мезенхімою. Мезенхіма між стінкою барабанної порожнини та сліпим кінцем зовнішнього слухового ходу утворює середній сполучнотканинний шар барабанної перетинки. Таким чином, барабанна перетинка утворюється за рахунок сполучення ентодерми І зябрової кишені з мезенхімою І зябрової дуги та ектодермою І зябрової щілини (тобто виникає з трьох зародкових листків).
Із маси мезенхіми І та ІІ зябрових дуг, яка лежить над майбутньою барабанною порожниною, диференціюються хрящові закладки слухових кісточок. По мірі свого розвитку барабанна порожнина збільшується в розмірі та охоплює слухові кісточки разом з мезенхімою, яка оточує їх. Таким чином, слухові кісточки опиняються зміщеними в барабанну порожнину. Мезенхіма, яка оточує слухові кісточки, дає початок сполучній тканині, яка розсмоктується після народження. Тільки після резорбції залишків сполучної тканини слухові кісточки набувають повної рухливості (порушення процесу резорбції може слугувати причиною вродженої глухоти).
За своїм розвитком слухові кісточки гомологічні кісткам підвіска та сполучення щелеп нижчих хребетних тварин з мозковим черепом. Молоточок і коваделко походять з І зябрової дуги та гомологічні сполучній (articulare) та квадратній (quadratum) кісткам, які утворюють сполучення верхньої та нижньої щелеп нижчих хребетних, включаючи рептилій. Сполучна кістка являє собою осифіковану базальну частину меккелевого хряща (сполучна поверхня примітивної нижньої щелепи). Квадратна кістка є кісткою дермального походження і являє собою сполучну поверхню верхньої щелепи. Стремінце у ссавців походить з найбільш дорсально розташованої частини ІІ зябрової дуги. У деяких риб ця кісточка називається гіомандибулярною кісткою та виконує функцію підвіска щелепного апарата до мозкового черепа. У амфібій гіомандибулярна кістка стала кіcткою в середньому вусі – columela. Ця кістка здійснює механічний зв’язок між барабанною перетинкою, яка знаходиться на поверхні голови, та внутрішнім вухом. У ссавців зазначені кістки, пов’язані зі щелепами, видозмінились у слухові кісточки і набули відповідних функцій.
М’язи слухових кісточок розвиваються з відповідних до розвитку кісточок зябрових дуг: м’яз-натягувач барабанної перетинки розвивається з І зябрової дуги; стремінцевий м’яз – із ІІ зябрової дуги.
Що стосується вушної раковини, то вона утворюється з мезенхіми, яка оточує першу (гіомандибулярну) зяброву щілину зародка людини. Упродовж другого місяця розвитку у зародка з’являється група з шести горбків; деякі з них виникають із тканин І зябрової дуги та розташовані попереду від І зябрової щілини і лежать вздовж задньої межі цієї щілини. Зрощення вказаних горбків та їх подальший розвиток призводять до утворення вушної раковини. Наявність великої кількості окремих центрів росту та складність формоутворюючого процесу обумовлює виразні індивідуальні розбіжності форми зовнішнього вуха.
Аномалії розвитку вуха
Аномалії внутрішнього вуха клінічно проявляються нейросенсорною приглухуватістю, глухотою або глухонімотою. Під терміном “приглухуватість” розуміють таке погіршення слуху, при якому хворому важко спілкуватися з людьми за допомогою розмовної мови. При глухонімоті дитина не чує розмовної мови, у зв’язку з чим вона не може навчатися нормальної розмовної мови (мовний апарат при цьому у неї не пошкоджений).
Приглухуватість, глухота та глухонімота можуть бути також наслідком такої вушної вади, як зрощення слухових кісточок – synotia, але в цьому випадку має місце порушення слуху за кондуктивним типом (тобто страждає апарат звукопроведення, а не звукосприйняття). Може також мати місце гіпоплазія (недорозвиток) усього середнього вуха (найрізноманітніші варіанти недорозвитку слухових кісточок, відсутність сполучення між ними, частіше між молоточком та коваделком), яка поєднується з гіпоплазією зовнішнього вуха; при аномаліях зовнішнього та середнього вуха внутрішнє вухо може виявитися цілковито нормальним, що пояснюється різницями в термінах і джерелах розвитку цих утворень.
Більшість різновидів уродженої глухоти зумовлена генетичними факторами, хоча чинники навколишнього середовища можуть також порушувати хід нормального розвитку внутрішнього і середнього вуха. Вірус краснухи, який вражає зародок на сьомому або восьмому тижні, здатен викликати значне ушкодження спірального органа. Існує думка, що причиною вродженої глухоти бувають також поліомієліт, еритробластоз плода, діабет, гіпотиреоз і токсоплазмоз.
Вади вушної раковини зустрічаються порівняно рідко, 1–2 випадки на 10 000, і як ізольовані порушення розвитку мають порівняно невелике клінічне значення. Однак досить часто ці вади розвитку бувають складовим компонентом синдромів – як спадкових, так і обумовлених зовнішніми факторами. Причому вигляд дисплазії вушної раковини має певне діагностичне значення. Так у дітей із синдромом Дауна вушні раковини зменшені, округлої форми, з широко звисаючим завитком; при синдромі Едвардса вушна раковина витягнута в горизонтальній площині, вушна часточка, а нерідко й козелок, відсутні.
Залежно від важкості та характеру морфологічних змін розрізняють:
1) вади вушної раковини з вродженою нестачею тканин, ступінь прояву якої коливається від мікротії (зменшена вушна раковина) до анотії (відсутність вушної раковини);
2) деформації, збільшення, серед яких розрізняють збільшену кількість вушних раковин (поліотію) та збільшення розмірів вушної раковини (макротію), розщілини вушної раковини, передвушні кісти та нориці.
Мікротія охоплює спектр дефектів зовнішнього вуха, починаючи від малих, але добре сформованих вушних раковин аж до їх повної відсутності (анотія). Поєднання кількох вад – мікротії, анотії та окулоаурикуловертебрального синдрому (геміфаціальна мікросомія) – трапляється у 20–40 % хворих дітей. У цих випадках черепно-лицеві дефекти можуть бути асиметричними. Оскільки певні структури зовнішнього та середнього вуха розвиваються з двох перших зябрових дуг, які утворюються, як недавно доведено, переважно клітинами нервового гребеня, ця клітинна популяція відіграє важливу роль у формуванні більшості вушних вад.
При синдромах множинних вроджених вад (особливо часто у випадках синдромів “котячого ока” та Гольденхара), а також як ізольована вада трапляються парааурикулярні папіломи (додаткові вушні раковини, “вушні придатки”) – фрагменти зовнішнього вуха, розташовані попереду вушної раковини.
Зустрічаються також передвушні (білявушні) нориці – ходи, що сліпо закінчуються, вивідні протоки яких розташовані біля основи висхідної частини завитка попереду козелка або вушної часточки. Закінчується нориця біля хряща слухового ходу, раковини вуха або біля завитка. Вкриті нориці епідермісом. Передвушні нориці часто супроводжують різні вади обличчя. Ізольовані форми зазвичай успадковуються за аутосомно-рецесивним типом.
Аномалії зовнішнього вуха трапляються при порушенні розвитку І та ІІ зябрових дуг – аномаладі І зябрової дуги та аномаладі І й ІІ зябрових дуг.
При аномаладі І зябрової дуги (першозябровий синдром, односторонній зяброво-лицевий дизостоз) відсутня тільки передня третина вушної раковини, в той час як задні дві третини збережені, хоч і можуть бути деформованими. Ця вада розвитку поєднується з одностороннім недорозвитком нижньої та верхньої щелепи, виличної кістки, жувальних м’язів, макростомією. Як видно з переліку змінених органів, страждають похідні І зябрової дуги. При аномаладі І та ІІ зябрових дуг (внутрішньоматковий лицевий некроз, некротична дисплазія, гемігнатія та мікротія) мають місце ті ж ознаки, що й при аномаладі І зябрової дуги, але вушна раковина та зовнішній слуховий хід практично відсутні (може залишатися тільки рудимент вушної часточки).