Органи кровотворення та імунної системи
В організмі людини існує система захисту від чужорідних клітин, інфекційних збудників та інших речовин, що мають антигенні властивості. Така функція опору організму до антигенів називається імунітетом (від латинського слова immunitas – звільнення від будь-чого), що забезпечує збереження сталості внутрішнього середовища і захисних реакцій організму. Наука про імунітет, що вивчає клітинні, генетичні і молекулярні механізми реагування організму на сторонні речовини, називається імунологією. Розділ імунології, який досліджує структурні основи захисних механізмів, називається імуноморфологією. Встановлення внутрісистемних та міжсистемних зв’язків імунних (лімфоїдних) органів, їх структурних змін при імунних процесах – основне завдання сучасної імуноморфології, що має клінічне спрямування.
Ще у 70-х роках ХХ століття впроваджено термін – імунна система, що об’єднує всі лімфоїдні органи, скупчення лімфоїдних клітин в інших органах. Особливістю цієї системи є її генералізація у всьому тілі, а її клітини постійно рециркулюють через кровоносне і лімфатичне русло. Імунні органи побудовані з лімфоїдної тканини, що представлена ретикулярною стромою, у якій розташовані лімфоїдні клітини – різноманітні субпопуляції Ті В-лімфоцитів, які забезпечують імунну відповідь на антигени за допомогою лімфоцитів-ефекторів. Плазмоцити, як В-ефекторні клітини, виробляють специфічні імуноглобуліни – антитіла – і забезпечують гуморальний імунітет. Т-кілери є Т-ефекторними клітинами і знешкоджують сторонні клітини, забезпечуючи клітинний імунітет.
Лімфатична система (systema lymphoideum), як частина імунної системи, має розгалужені в органах і тканинах лімфатичні капіляри, лімфокапілярні сітки, лімфатичні судини, стовбури і протоки. На шляхах току лімфи розташовані численні лімфатичні вузли, що є біологічними “фільтрами” для лімфи, яка протікає через них.
Принципи етапності лімфовідтоку були вперше сформульовані засновником української лімфології київським професором Ф. А. Стефанісом. Відомий сучасний лімфолог академік М. Р. Сапін у книзі
«Внеорганные пути транспорта лимфы» (1982) підкреслив, що «четкое определение поэтапного оттока лимфы было дано Ф. А. Стефанисом», а «значение этапности оттока лимфы от органов имеет важное значение в клинике при определении путей распространения метастазов опухолей и воспалительных процессов». У науковій роботі «Лимфатические сосуды желудка человека» (1900) Ф. А. Стефаніс писав: «Первым этапом будет путь от места происхождения данного лимфатического сосуда и до первого встреченого им узла, а первым этапным узлом – этот узел; выносящий сосуд первого этапного узла будет представлять собой второй этап, а узел, в который он впадает, – второй этапный узел и т. д.».
Нині особливу увагу приділено мікролімфоциркуляції в нормі і при різних патологічних процесах в організмі. За останні роки в клініку впроваджено методи хірургічного реконструювання лімфатичних і венозних судин – створення штучних лімфовенозних анастомозів для лікування важких набряків, причиною яких є структурні зміни лімфатичних судин. Широко застосовуються способи дренування лімфатичних проток та стовбурів з метою детоксикації організму; ендолімфатичного введення лікарських засобів для корекції певних порушень в організмі; введення світловодів лазерів для ендолімфатичного опромінення лімфи тощо.
У нашому підручнику анатомічні терміни відповідають „Міжнародній анатомічній номенклатурі – українському стандарту” (Київ, 2001), тому читач побачить певні термінологічні відмінності від підручників попередніх років видання, у яких органи і судини лімфатичної системи подають в розділі “Органи кровотворення та імунної системи”. Зокрема, в них використовують терміни: “Імунна система, центральні і периферійні лімфоїдні органи, лімфатичні вузлики”, а за українським стандартом, який доповнено і затверджено на IV Національному конгресі АГЕТ України в 2006 році, вони мають таку назву: “Лімфатична система, первинні і вторинні лімфоїдні органи, лімфоїдні вузлики”. Одним із структурних компонентів вторинних лімфоїдних органів є лімфоїдний вузлик, який у підручниках ще називають лімфатичним вузликом, або лімфоїдним фолікулом.
Лімфатична система (systema lymphoideum), морфологічно і функціонально об’єднана з кровоносною системою, є частиною єдиної судинної та імунної системи. Цей термін походить від латинського слова lympha – чиста вода джерела.
Функція лімфатичної системи. Через лімфатичні капіляри всмоктується міжклітинна рідина, з якою у лімфатичні судини потрапляють продукти клітинного обміну, лімфоцити, часом навіть еритроцити, сторонні речовини, зокрема, різноманітні антигени – фрагменти відмерлих клітин і тканинних елементів, клітини-мутанти, мікроорганізми, віруси тощо. У лімфатичних вузлах лімфа „фільтрується” і очищається від цих продуктів. Макрофаги фагоцитують антигени, „переробляють” їх і передають імунну інформацію лімфоцитам. Так запускається процес антигензалежної проліферації та диференціації субпопуляцій Ті В-лімфоцитів, формується конкретна імунна відповідь. Отже, лімфатична система виконує захисну функцію.
Лімфа (lympha) утворюється внаслідок всмоктування міжклітинної рідини в капіляри лімфатичної системи. Це безбарвна прозора рідина, яка подібна за біохімічним складом до плазми крові. Основними клітинними елементами лімфи (96–98 %) є лімфоцити. У лімфатичній системі дорослої людини циркулює приблизно два літри лімфи. Якщо людина з’їла багато жирної їжі, то лімфа, яка відтікає від кишки, стає білою як молоко (молочний сік, chylus). Грецьке слово chylus означає “штучно приготовлений сік”. Цим терміном називали лімфу кишкових лімфатичних судин.
Відкриття лімфатичних судин пов’язане з іменем італійського анатома XVII століття Гаспаро Азеллі (1581–1626). У 1622 році, досліджуючи рухи діафрагми у живої собаки, він виявив у брижі тонкої кишки судини, які були наповнені білуватим молокоподібним вмістом. З’ясувалося, що перед експериментом собаку нагодували жирною їжею. Азеллі назвав ці судини молочними і вважав, що вони несуть “білу кров”, або молочний сік (хілус) у печінку. Згодом, після смерті Азеллі, було встановлено, що в цих судинах тече лімфа у венозну систему.
Функціональне значення грудної протоки, відомої під назвою „біла вена”, було з’ясоване студентом Паризького університету Ж. Пеке (1622–1674). Експериментальним шляхом у 1647 році він встановив, що лімфа з органів черевної порожнини рухається до грудної протоки, яка впадає у вени шиї.
У людини грудну протоку описали О. Рудбек (1651) і Ф. Бартоліні (1652). Клапани в лімфатичних судинах описав Ф. Рюїл (1655).
Завдяки дослідженням відомих анатомів П. Москаньї (1787), Ф. Саппея (1810–1896), Г. М. Йосифова (1870–1933), Г. Рув’єра (1932), академіків Д. А. Жданова (1908–1972) і М. Р. Сапіна, але особливо завдячуючи роботам українських анатомів-лімфологів київської школи Ф. А. Стефаніса (1865–1917), М. С. Спірова (1896–1972), А. А. Сушка (1899–1970),
О. І. Свиридова (1900–1973) було створене і розвинуте вчення про лімфатичну систему.
ОРГАНИ КРОВОТВОРЕННЯ ТА ІМУННОЇ СИСТЕМИ
Класифікація лімфоїдних (імунних) органів
Лімфатична система складається з первинних і вторинних лімфоїдних (імунних) органів та лімфоносних судин.
До первинних лімфоїдних органів (organa lymphoidea primaria) належать:
– червоний кістковий мозок (medulla ossium rubra), який є аналогом бурси Фабриціуса у птахів і в якому відбувається антигеннезалежна проліферація і диференціація субпопуляцій В-лімфоцитів;
– загруднинна залоза, або тимус (thymus), у якій відбувається антигеннезалежна проліферація і диференціація субпопуляцій Т-лімфоцитів.
Субпопуляції Ті В-лімфоцитів потім “заселяють” Ті В-залежні зони у вторинних лімфоїдних органах, де формується конкретна імунна відповідь на антигени.
До вторинних лімфоїдних органів (organa lymphoidea secundaria) належать:
– лімфатичні вузли (nodi lymphatici; nodi lymphoidei; lymphonodi);
– мигдалики (tonsillae), які містяться в ділянці
глотки. Вони утворюють лімфатичне (лімфоїдне) кільце глотки (anulus lymphoideus pharyngis) із шести мигдаликів: одного язикового мигдалика (tonsilla lingualis), парного піднебінного мигдалика (tonsilla palatinа), парного трубного мигдалика (tonsilla tubaria) і одного глоткового мигдалика (tonsilla pharyngea). Ці органи складаються з лімфоїдної тканини, що утворює численні лімфоїдні вузлики (noduli lymphoidei);
– скупчення лімфоїдної тканини в стінках органів травної і дихальної систем, сечовидільних шляхів у вигляді одиноких лімфоїдних вузликів (noduli lymphoidei solitarii), діаметром до 1,5–2 мм. Якщо скупчується п’ять і більше таких вузликів, то утворюється скупчений лімфоїдний вузлик (nodulus lymphoideus aggregaticus). Його ще називають лімфоїдною бляшкою, або бляшкою Пейєра. Таких бляшок багато у стінці клубової кишки, їх довжина становить від 0,2 до 15 см, а ширина – 0,2–1,5 см;
–червоподібнийвідросток(appendixvermiformis), у слизовій оболонці і в підслизовій основі якого розташовано до 550 одиноких лімфоїдних вузликів різної форми та розмірів (від 0,2 до 1,2 мм);
– селезінка (splen, lien), зокрема, її біла пульпа, що складається з лімфоїдних вузликів і лімфоїдних періартеріальних піхв.
У вторинних лімфоїдних органах відбувається антигензалежна проліферація і диференціація субпопуляцій Ті В-лімфоцитів, які формують конкретну імунну відповідь.
Органи кровотворення та імунної системи тісно пов’язані між собою спільністю походження будови і функцій. Родоначальником усіх видів клітин крові та імунної системи є поліпотентні стовбурові клітини червоного кісткового мозку, тому таку клітину називають стовбуровою кровотворною клітиною (СКК). У червоному кістковому мозку на 105 ядерних клітин припадає приблизно 50 СКК. В організмі дорослої людини є в середньому 5 × 105 стовбурових кровотворних клітин, третина з яких перебуває в мітотичному циклі. Стовбурові клітини здатні до численного мітотичного поділу, зокрема стовбурові клітини кісткового мозку можуть поділятися до 100 разів. При мітотичному поділі з двох дочірніх клітин одна клітина залишається стовбуровою, а друга диференціюється. Однак стовбурові клітини детерміновані, тобто вони диференціюються лише у визначеному напрямку і передають свої ознаки наступним клітинним поколінням. Ця детермінація обумовлена генетично.
Стовбурові клітини здатні утворювати колонії, кожна з них є клоном, що виник з однієї клітини. Поліпотентна стовбурова клітина кісткового мозку називається колонієутворюючою одиницею (КУО). Незначна кількість колонієтвірних клітин циркулює в крові. У червоному кістковому мозку, у його гемоцитопоетичній (мієлоїдній) тканині, зі стовбурових клітин утворюються клітини-попередниці, з яких, шляхом мітотичного поділу і диференціації, виникають формені елементи крові. Зі стовбурових клітин у кістковому мозку утворюється також лімфоцитопоетична тканина, що дає початок популяції лімфоцитів – робочим клітинам імунної системи.
Напрямок диференціації колонієтвірних клітин визначається після того, як клітина-попередниця вступає на визначений шлях диференціації, тобто стає комітивною. Таким клітинам необхідні специфічні глікопротеїнові гормони, що контролюють їх виживання і диференціацію через вплив на активність генів. Зараз відомі шість класів диференціації клітин крові та імунної системи під впливом специфічних глікопротеїнових чинників (див. підручники з гістології та імунології).
Кровотворення в людини починається наприкінці другого – початку третього тижнів ембріогенезу в стінці жовткового мішка – це зародковий гемоцитопоез, де вперше утворюються кров’яні острівці. У цих острівцях з клітин мезенхіми утворюються стовбурові клітини, які в просвіті судин (інтраваскулярно) диференціюються на клітини крові. Після редукції жовткового мішка, починаючи з 7–8-го тижня розвитку зародка, кровотворення продовжується в печінці – печінковий гемопоез. Зі стовбурових клітин, що надійшли в печінку із судин (екстраваскулярно), також утворюються клітини крові. Кровотворення в печінці продовжується до кінця внутрішньоутробного періоду розвитку плода.
Кровотворення в кістковому мозку (кістковомозковий гемопоез), який закладається на другому місяці ембріогенезу, починається на 12-му тижні розвитку зародка і триває протягом усього життя людини. Отже, кровотворним органом у людини після народження є червоний кістковий мозок (medulla ossium rubra).
Зі стовбурових клітин екстраваскулярно розвиваються клітини крові : еритроцити (еритропоез), гранулоцити (гранулопоез) і тромбоцити (тромбопоез), а також формуються моноцити, що належать до макрофагальної системи (моноцитопоез) і клітини імунної системи – В-лімфоцити (лімфоцитопоез). Стовбурові клітини виселяються також з кісткового мозку в загруднинну залозу (тимус), де вони диференціюються в Т-лімфоцити.
Імунна система об’єднує органи і тканини, що забезпечують захист організму від генетично чужорідних клітин та речовин (антигенів), які надходять іззовні чи утворюються в організмі. До органів імунної системи (лімфоїдних органів) належать всі органи, які беруть участь в утворенні клітин, що здійснюють захисні реакції в організмі, а саме: різні субпопуляції Ті В-лімфоцитів, зокрема плазмоцитів. Імунні органи побудовані з лімфоїдної тканини, що розміщена в ретикулярній стромі. У її петлях містяться клітини лімфоїдного ряду: лімфоцити різного ступеня зрілості (бласти, великі, середні і малі лімфоцити), молоді і зрілі плазматичні клітини (плазмобласти, плазмоцити).
Стовбурові клітини, що надходять із червоного кісткового мозку в кров, уже на 7–8-й тиждень ембріогенезу заселяють загруднинну залозу (тимус), де з них утворюються тимусзалежні лімфоцити – Т-лімфоцити. Тобто в цьому органі відбувається антигеннезалежна диференціація і проліферація субпопуляцій Т-лімфоцитів. У червоному кістковому мозку із стовбурових клітин утворюються В-лімфоцити – бурсозалежні лімфоцити. Таку назву ці клітини отримали тому, що зараз червоний кістковий мозок вважають аналогом бурси (сумки) Фабриціуса – клітинного скупчення в стінці клоаки птахів. Обидві популяції Ті В-лімфоцитів з течією крові надходять із загруднинної залози (тимуса) і червоного кісткового мозку у вторинні лімфоїдні органи імунної системи. Необхідно підкреслити, що всі субпопуляції Ті В-лімфоцитів утворюються з поліпотентних стовбурових клітин червоного кісткового мозку.
Т-лімфоцити заселяють тимусзалежні зони, зокрема в лімфатичних вузлах (паракортикальна зона), селезінці (лімфоїдні періартеріальні муфти). В-лімфоцити, які є попередниками плазмоцитів, що виробляють специфічні білки – імуноглобуліни (антитіла), і лімфоцитів з підвищеною активністю, надходять у В-зони (бурсозалежні зони) лімфатичних вузлів (лімфоїдні вузлики, мозкові тяжі) і селезінки (лімфоїдні вузлики, окрім їхньої періартеріальної зони) та інших лімфоїдних органів.
У вторинних лімфоїдних органах відбувається антигензалежна диференціація і проліферація субпопуляцій Ті В-лімфоцитів, які формують конкретну імунну відповідь організму.
Отже, вторинні лімфоїдні органи виробляють імунокомпетентні клітини, зокрема лімфоцити, включають їх в імунний процес, розпізнають, знищують чужорідні речовини, що потрапили в організм або утворилися в ньому і містять у собі “ознаки генетично чужорідної інформації”. Такі чужорідні речовини називаються антигенами. При проникненні в організм антигенів у вторинних лімфоїдних органах проти них утворюються нейтралізуючі речовини – антитіла, що є імуноглобулінами, забезпечуючи гуморальний імунітет. Синтезують антитіла плазматичні клітини (плазмоцити), що є ефекторними В-лімфоцитами. Антитіла виділяються в кров і у секрет залоз, з’єднуються з відповідними антигенами і нейтралізують їх, згодом ці комплекси захоплюють фагоцити. Ефекторні Т-лімфоцити – Т-кілери забезпечують в основному клітинний імунітет. Т-кілери знищують чужорідні, а також змінені власні клітини.
Дослідженнями останніх років з’ясовано, що імунна система, як і інші системи організму, наділена всіма характеристиками великих систем: наявність цілі, багатокомпонентність, багатозв’язковість, багаторівнева ієрархічна структура, числені форми управління, складність поведінки, стабільність, надійність. Відмінності імунної системи від інших зумовлюють цілі її функціонування. Фізіологи сучасності вбачають їх у звільненні внутрішнього середовища організму від продуктів з чужорідною генетичною інформацією, означених як антигени. У ширшому понятті імунна система реалізує захист організму від генетично чужорідних клітин і речовин, які надходять іззовні чи утворюються безпосередньо в організмі. Беззаперечним є уявлення про імунологічні процеси як про інтегративну ділянку гомеостазу. Пластичність останнього формується при взаємодії організму й оточуючого середовища. Під впливом певних стимулів в організмі відбувається ряд молекулярних і клітинних перетворень, що контролюються на різних рівнях регуляторними реакціями. В основу останніх покладено принцип зворотного зв’язку. Поряд із цим, імунологічний нагляд деякі науковці розглядають як генетично контрольований і регульований взаємозв’язок специфічних імунологічних і неспецифічних біологічних захисних реакцій. Саме в цьому контексті імунологічний контроль представляється у новій якості як імунобіологічний нагляд, що об’єднує в собі імунологічні і біологічні наглядові механізми та ефектори.
Як зазначалося вище, імунна система включає первинні (тимус, червоний кістковий мозок) і вторинні органи (лімфатичні вузли, селезінка, мигдалики, одинокі лімфоїдні вузлики слизових оболонок, скупчені лімфоїдні вузлики червоподібного відростка та клубової кишки). Встановлено, що після завершення розвитку в первинних органах імунної системи лімфоцити покидають їх і заселяють вторинні лімфоїдні органи. Організація вторинних органів імунної системи повністю підпорядкована їх функціональному призначенню. Суть останнього полягає в ідентифікації та елімінації чужорідних субстанцій. Для виконання такої складної роботи вкрай необхідне строго впорядковане заселення вторинного відділу імунної системи лімфоїдними клітинами, а також підтримання клітинних популяцій у динамічному стані.
Лімфоцити вторинних лімфоїдних органів не затримуються в них надовго завдяки реалізації двох процесів, що забезпечують їх невпинну круговерть, – поновлення та рециркуляції. Рециркуляцію лімфоцитів розглядають як процес безперервного переміщення цих клітин із органів у лімфота кровоплин і їх наступне повернення (феномен хомінгу). Мета рециркуляції полягає в постійному патрулюванні тканин організму імунокомпетентними лімфоцитами, ефективному розпізнаванні чужорідних і власних (ендогенних) антигенів, наділенні органів імунної системи інформацією про наявні у тканинах антигени. Завдяки рециркуляції формується єдиний пул зрілих лімфоцитів і не допускаються навіть випадкові “викривлення” клонального репертуару в різновіддалених ділянках організму. Міграція є важливим процесом також і для функціонування центральної ланки імунної системи. У цій ланці відповідно при реалізації певних закономірностей і сигналів до міграційних процесів залучаються стовбурові кровотворні клітини, попередники Т-лімфоцитів, дозріваючі Ві Т-лімфоцити.
Із загальної кількості лімфоцитів, маса яких у тілі дорослої людини дорівнює приблизно 1500 г (6 • 1012 клітин), у периферійній крові (окрім кровотворних і лімфоїдних органів) циркулює тільки 0,2 % лімфоцитів – приблизно 3 г, тобто 10 • 129 клітин. Інші лімфоцити містяться в лімфоїдній тканині органів імунної системи – 100 г, у червоному кістковому мозку – 100 г, в лімфі та інших тканинах – 1300 г. У 1 мкл лімфи грудної протоки міститься від 2000 до 20000 лімфоцитів, а в 1 мкл периферійної лімфи, що відтікає від лімфокапілярних сіток до лімфатичних вузлів, є в середньому тільки 200 лімфоцитів.
У немовлят загальна маса лімфоцитів дорівнює в середньому 150 г, з них тільки 0,3 % циркулює в крові. Потім кількість лімфоцитів швидко наростає, у дітей віком від 6 місяців до 6 років їх маса вже становить приблизно 650 г. У 15-річних дітей масса всіх лімфоцитів збільшується до 1250 г, а в периферійній крові є тільки 0,2 % від усієї маси цих клітин.
Лімфоцити є дуже рухливими клітинами, їхній діаметр коливається від 5 мкм до 18 мкм. Більшість циркулюючих лімфоцитів мають діаметр 5–7 мкм – це малі лімфоцити. Приблизно 10 % складають середні лімфоцити діаметром 8–10 мкм. Великі лімфоцити (лімфобласти) мають діаметр 11–18 мкм, вони переважно містяться в центрах розмноження (світлих центрах) лімфоїдних вузликів лімфатичних вузлів і селезінки. У нормі вони не циркулюють у крові і присутні в лімфі. У малих лімфоцитах майже весь об’єм клітин займає велике кругле ядро. У середніх лімфоцитах вузький обідок цитоплазми товщиною 2–3 мкм оточує велике ядро. Для великих лімфоцитів характерне велике кругле світле ядро, гомогенно заповнене еухроматином (див. підручники з гістології та імунології). Малі лімфоцити є основними імунокомпетентними клітинами. Вважають, що середні лімфоцити є початковою стадією диференціації В-лімфоцитів у плазматичні клітини (плазмоцити).
У малих і середніх лімфоцитах ядра розташовані в центрі клітин, вони мають незначні інвагінації. В ядрах переважає конденсований хроматин – гетерохроматин, ядерце чітко визначається при електронномікроскопічному дослідженні. У цитоплазмі цих клітин багато вільних рибосом, що зумовлює її інтенсивну базофілію, відносно мало мітохондрій і коротких цистерн гранулярної ендоплазматичної сітки, комплекс Гольджі слабо виражений, але є центросома. Приблизно у 10 % циркулюючих лімфоцитів у цитоплазмі міститься невелика кількість азурофільних гранул.
При світловій мікроскопії розрізнити за морфологічними ознаками Ті В-лімфоцити неможливо. Їх субпопуляції можна ідентифікувати тільки за допомогою імуногістохімічних методів. При електронній мікроскопії видно, що лімфоцити мають на поверхні ультрамікроскопічні вирости – мікроворсинки. Мікроворсинки містять своєрідні білки-рецептори (чуттєві апарати), що розпізнають антигени. Антигени викликають в організмі імунні реакції, що супроводжуються утворенням антитіл В-ефекторними лімфоцитами – плазматичними клітинами (плазмоцитами). У вторинних лімфоїдних органах плазмоцити контактують з іншими лімфоцитами, макрофагами і ретикулярними клітинами. Щільність мікроворсинок на поверхні В-лімфоцитів у 100–200 разів більша, ніж на поверхні Т-лімфоцитів. На поверхні лімфоцитів є специфічні білки – маркери, що є різними в Ті В-клітинах. За тими маркерами імуногістохімічними методами визначають різні субпопуляції Ті В-лімфоцитів.
Структура антитіл, генетичні механізми їх утворення і різноманіття описують в курсах молекулярної біології та генетики. Ми обмежимося лише короткими повідомленнями, що необхідні для розуміння морфофункціональної організації імунної системи. Основною властивістю клітин імунної системи (імунокомпетентних клітин) є їх здатність взаємодіяти з численними антигенами. Доцільно нагадати, що діяльність людства за останні десятиріччя призвела до значного зростання різновидів антигенів, а їх кількість продовжує катастрофічно збільшуватись, викликаючи різноманітні патологічні стани в організмі людей. У даний час вважають, що кожен В-лімфоцит програмується в мієлоїдній тканині червоного кісткового мозку, а кожен Т-лімфоцит – у кірковій речовині загруднинної залози (тимуса). У результаті цього лімфоцити “індивідуально” запрограмовані на взаємодію з визначеними антигенами і в цьому сенсі є високоспеціалізованими клітинами. У процесі програмування на цитолемі лімфоцитів утворюються специфічні білки-рецептори, які є комплементарними до визначеного антигена. Зв’язування певного антигена з комплементарним рецептором викликає ряд послідовних реакцій, що призводять до проліферації даної клітини й утворення великої кількості нових лімфоцитів, які реагують тільки з даним антигеном.
Отже, однією з найважливіших властивостей імунної системи є імунна пам’ять. У результаті першої зустрічі запрограмованого лімфоцита з визначеним антигеном утворюються дві категорії клітин: ефекторні клітини, що негайно виконують специфічну імунну функцію – секретують відповідні антитіла чи знешкоджують чужорідні клітини, і клітини пам’яті, що довго циркулюють в організмі. При повторному надходженні даного антигена в організм Т і В-лімфоцити пам’яті швидко перетворюються в лімфоцити-ефектори, що вступають в реакцію з антигеном. При кожному мітотичному поділі запрограмованого лімфоцита після його зустрічі з антигеном кількість клітин пам’яті збільшується. Отже, у вторинних лімфоїдних органах імунної системи присутні три функціональних типи лімфоцитів: запрограмовані Ті В-лімфоцити, Ті В-ефектори, Ті В-клітини пам’яті. На цитолемі запрограмованого В-лімфоцита містяться ділянки розпізнавання антигена – поверхневі рецептори, що представлені вбудованими в цитоплазматичну мембрану молекулами специфічного імуноглобуліну, які розпізнають конкретний антиген.
При контакт із антигеном запрограмовані Т-лімфоцити активуються, збільшуються за розмірами, мітотично поділяються. Кожна з дочірніх клітин диференціюється у визначену субпопуляцію. Розрізняють п’ять субпопуляцій Т-лімфоцитів, кожна з яких обумовлює визначену відповідь. Одна з них – Т-кілери (від англійського to kill – убивати). При зустрічі зі специфічною чужорідною клітиною відросток Т-кілера короткочасно контактує з цитолемою чужорідної клітини – клітини-мішені, руйнує (розчиняє) її цитолему, що веде до підвищення проникності клітинної мембрани клітини-мішені. У результаті цього з клітини виходять іони калію, а в клітину входять іони натрію і вода. Клітина набухає і руйнується (лізується). Т-супресори гальмують імунну відповідь В-лімфоцитів та інших Т-лімфоцитів на антигени за допомогою утворених ними розчинних факторів. Т-ампліфікатори є джерелом підтримки популяції Т-лімфоцитів і не рециркулюють. Це нечисленна популяція короткоживучих клітин, що містяться в загруднинній залозі (тимусі) і селезінці.
Як відомо, В-лімфоцити є попередниками плазматичних клітин, що синтезують і виділяють імуноглобуліни – антитіла. Однак для здійснення імунної відповіді на антиген В-лімфоцитам необхідна їх кооперація з іншими Т-лімфоцитами – Т-хелперами (від англійського to help – допомагати), що виділяють Т-хелперний фактор у відповідь на зв’язування антигена зі специфічним рецептором у мембрані Т-лімфоцитів. В-лімфоцити активуються лише при наявності Т-хелперного фактора. Тобто, для здійснення імунної відповіді конче необхідна кооперація Ті В-лімфоцитів. Окрім того, в цьому процесі обов’язково беруть участь макрофаги. Отже, для ефективної імунної відповіді необхідне утворення триклітинної системи – Т-лімфоцити, В-лімфоцити і макрофаги.
Т-ефектори, які утворилися після взаємодії Т-лімфоцитів з антигеном, виробляють і виділяють біологічно активні речовини – лімфокіни. Серед них є такі, що впливають на макрофаги: одні пригнічують міграцію макрофагів, другі їх активують, а інші викликають агрегацію макрофагів. Окрім того, синтезуються чинники переносу, хемотаксису, мітогенні, які стимулюють або пригнічують утворення антитіл, специфічний і неспецифічний фактори, що гальмують проліферацію клітин, тощо. Важливу роль у противірусному захисті організму відіграє інтерферон, який синтезується Т-лімфоцитами. Т-лімфоцити беруть участь у реакціях відторгнення пересаджених трансплантатів.
Однією з найважливіших особливостей Т-лімфоцитів є їх здатність утворювати “розетки” з еритроцитами барана, що має діагностичне значення. У цитоплазмі Т-лімфоцитів багато лісозом, а кисла фосфатаза, на відміну від В-лімфоцитів, має високу активність. Більшість Т-лімфоцитів швидко рециркулює, тривалість життя таких клітин досягає 4–6 місяців. В-лімфоцити рециркулюють повільніше, а живуть декілька тижнів. Т-лімфоцити не мають рецепторів до комплементу і комплексів “антиген – антитіло”, а у В-лімфоцитів такі рецептори наявні.
Середній вміст популяцій Т і В-лімфоцитів у лімфоїдних (імунних) органах та периферійній крові представлений у таблиці 24.
Лімфоїдні (імунні) органи мають характерні закономірності розвитку і будови в пренатальному і постнатальному онтогенезі:
- Робочою паренхімою всіх первинних і вторинних лімфоїдних (імунних) органів є лімфоїдна тканина, що складається із субпопуляцій Ті В-лімфоцитів, в ній обов’язково присутні макрофаги.
- Усі первинні і вторинні лімфоїдні (імунні) органи закладаються на початкових етапах пренатального онтогенезу: кістковий мозок і загруднинна залоза (тимус) починають формуватися на 4–5 тижні ембріогенезу; селезінка – на 5–6 тижні; лімфатичні вузли – на 7–8 тижні; піднебінні і глоткові мигдалики – на 9–14 тижні; скупчені лімфоїдні вузлики (бляшки) тонкої кишки і лімфоїдні вузлики червоподібного відростка – на 14–16 тижні; одинокі лімфоїдні вузлики слизових оболонок внутрішніх органів – на 16–18 тижні; язиковий мигдалик – на 24–25 тижні; трубні мигдалики – на 28–32 тижні.
- До моменту народження всі первинні і вторинні лімфоїдні (імунні) органи сформовані та функціонально зрілі. У немовлят вони вже виконують захисну функцію. Червоний кістковий мозок, що містить стовбурові клітини, мієлоїдну і лімфоїдну тканини, ще до народження дитини міститься у всіх кістковомозкових порожнинах. Загруднинна залоза (тимус) немовлят має таку ж відносну масу, як у дітей і підлітків, і складає 0,3 % від маси тіла. Лімфоїдні вузлики у вторинних лімфоїдних (імунних) органах, зокрема, в піднебінних мигдаликах і червоподібному відростку, вже сформовані у плода останніх місяців розвитку, що є ознакою зрілості органів імуногенезу.
- Усі первинні і вторинні лімфоїдні (імунні) органи досягають максимального розвитку (маса, розміри, кількість лімфоїдних вузликів) у дитячому віці і у підлітків.
- Для всіх первинних і вторинних лімфоїдних (імунних) органів характерна рання вікова інволюція, починаючи з підліткового та юнацького віку. У цих органах поступово зменшується кількість лімфоїдних вузликів, у них зникають центри розмноження (світлі центри), зменшується загальна кількість лімфоїдної тканини. Замість неї поступово збільшується кількість жирової й сполучної тканини.
- Первинні лімфоїдні (імунні) органи розташовані в організмі в добре захищених від зовнішніх впливів місцях: кістковий мозок розміщений у кістковомозкових порожнинах; загруднинна залоза (тимус) – у грудній порожнині за грудниною. Лімфоїдна тканина має специфічне мікрооточення: у кістковому мозку таким середовищем є мієлоїдна тканина, а в загруднинній залозі (тимусі) – епітеліальна тканина (епітеліоретикулоцити).
- Вторинні лімфоїдні (імунні) органи розташовані в місцях найімовірнішого проникнення в організм антигенів – генетично чужорідних речовин і клітин, або на шляхах їх розповсюдження в організмі. Наприклад, лімфатичне (лімфоїдне) кільце глотки (кільце Пирогова – Вальдейєра) оточує вхід у глотку із носової і ротової порожнин. У слизовій оболонці та підслизовій основі органів травної, дихальної і сечової систем розташовані численні скупчення лімфоїдної тканини у вигляді одиноких і скупчених лімфоїдних вузликів. Лімфатичні вузли розміщені на шляхах руху лімфи від органів і тканин. Через селезінку протікає кров із артеріальної системи у венозну – це орган, що „контролює” антигенний статус крові.
- У пренатальному і постнатальному онтогенезі у вторинних лімфоїдних органах постійно відбуваються процеси диференціації лімфоїдної тканини від дифузно розташованих клітин лімфоїдного ряду до лімфоїдних вузликів з центром розмноження, де відбувається антигензалежна проліферація і диференціація субпопуляцій Ті В-лімфоцитів. При антигенних впливах кількість вузликів із центром розмноження значно зростає. Спочатку утворюються скупчення лімфоїдних клітин без чітких границь, яке називають дифузною лімфоїдною тканиною або передвузликовою стадією формування вторинних лімфоїдних (імунних) органів. Такі дифузні скупчення лімфоїдної тканини можна спостерігати у внутрішньоутробному періоді розвитку людини на місці майбутніх мигдаликів, скупчених лімфоїдних вузликів, лімфатичних вузлів, а також у постнатальному онтогенезі в стінках стравоходу, дихальних і сечовидільних шляхів, де немає постійного і тривалого антигенного впливу на слизову оболонку, на відміну від шлунка, тонкої і товстої кишок, де їжа затримується досить довго.
Надалі дрібні дифузні скупчення лімфоїдної тканини ще більше ущільнюються, утворюючи передвузлики, які вже мають чіткі межі. У великих скупченнях лімфоїдної тканини (мигдаликах, скупчених лімфоїдних вузликах, лімфатичних вузлах) клітини лімфоїдного ряду в певних місцях ще більше ущільнюються, утворюючи лімфоїдні вузлики. Такі лімфоїдні вузлики утворюються в лімфоїдних (імунних) органах незадовго до народження дитини або в перші дні після народження. Наявність лімфоїдних вузликів у лімфоїдній тканині мигдаликів, лімфатичних вузлів, селезінки, у слизовій оболонці шлунка, кишки тощо є морфологічною ознакою зрілості органів імунної системи, їхньої готовності виконувати захисні функції у повному обсязі.
Найвищим ступенем диференціації органів імунної системи є поява у лімфоїдних вузликах центрів розмноження (світлих, або гермінативних центрів). Такі центри розмноження виникають у лімфоїдних вузликах при довготривалій чи сильній дії антигенів. Інтенсивне утворення центрів розмноження в лімфоїдних вузликах спостерігається вже у дітей грудного віку. У дітей перших трьох років життя до 70 % лімфоїдних вузликів у стінках тонкої кишки мають центри розмноження. Наявність центрів розмноження свідчить, з одного боку, про вплив на організм сильних і різноманітних антигенних чинників зовнішнього середовища, а з іншого боку – про велику функціональну активність захисних сил організму.
У лімфоїдних вузликах без центрів розмноження – первинних лімфоїдних вузликах – лімфоїдні клітини (переважно малі лімфоцити) розташовані щільно і відносно рівномірно. У лімфоїдних вузликах з центром розмноження – вторинних лімфоїдних вузликах – периферійна частина вузлика, його мантія, що розташована навколо центрів розмноження, складається з компактно розміщених малих і середніх лімфоцитів. Центри розмноження, де відбувається утворення нових лімфоцитів, містять багато лімфобластів і великих лімфоцитів, а також клітин на різних стадіях мітотичного поділу. Щільність клітин у них менша, ніж у мантії вузлика, тому на гістологічних препаратах центри розмноження видаються світлішими, звідси і їх друга назва – світлі центри. Починаючи з підліткового віку, кількість і розміри лімфоїдних вузликів поступово зменшуються, зникають центри розмноження. Після 40–60 років на місці більшості лімфоїдних вузликів залишається дифузна лімфоїдна тканина, що з віком частково замінюється жировою тканиною.
ПЕРВИННІ ЛІМФОЇДНІ (ІМУННІ) ОРГАНИ
Кістковий мозок
Кістковий мозок (medulla ossium) – це орган кровотворення і первинний лімфоїдний (імунний) орган (аналог бурси Фабриціуса у птахів), у якому відбувається антигеннезалежна проліферація і диференціація субпопуляцій В-лімфоцитів із стовбурових клітин. Виділяють червоний кістковий мозок (medulla ossium rubra), розміщений у комірках губчастої речовини плоских і коротких кісток, а також в епіфізах довгих кісток, і жовтий кістковий мозок (medulla ossium flava), розташований у кістковомозкових порожнинах довгих трубчастих кісток. У дорослої людини маса кісткового мозку становить 2,5– 3 кг (4,5–4,7 % від маси тіла), половина з якого – червоний кістковий мозок. Строму червоного кісткового мозку утворюють ретикулярні клітини і ретикулярні волокна. У комірочках строми розташовані гемоцитопоетичні (мієлоїдна тканина) і лімфоїдні (лімфоїдна тканина) елементи на різних стадіях розвитку, а саме: молоді і дозрілі клітини крові, плазмоцити, лімфоцити, лаброцити, ліпоцити, остеогенні клітини, макрофагоцити. Там обов’язково є стовбурові клітини – попередники всіх клітин крові, зокрема, лімфоцитів.
Ретикулярні клітини (інтердигітуючі клітини) не тільки є стромальними підтримуючими клітинами, але й мають фагоцитарні властивості і надають лімфоцитам інформацію про антиген. Ретикулярні клітини поліморфні – від зірчастої за формою клітини з численними відростками до веретеноподібної. Тонкі пучки ретикулярних волокон розміщені поблизу ретикулярних клітин, але вони не інвагінують у плазматичну мембрану, як у селезінці чи лімфатичних вузлах. Ретикулярні волокна переходять на межі кістковомозкової порожнини в пухкий шар колагенових волокон.
Червоний кістковий мозок розташований у вигляді тяжів (шнурів) циліндричної форми навколо артеріол. Тяжі розмежовані між собою великими кровоносними капілярами – синусоїдними капілярами (vasa capillaria sinusoidea), стінка яких утворена з ендотеліоцитів, вкритих ззовні тонким шаром колагенових волокон. Кровоносні судини кісткового мозку є гілками артерій, які живлять кістку. Характерною особливістю цих артерій є те, що в їх середній оболонці мало гладком’язових клітин, а зовнішня оболонка (адвентиція) дуже тонка. Від артерій відходять численні артеріоли, що розгалужуються на синусоїдні капіляри. Між ендотеліоцитами синусоїдних капілярів проникають у кровоносне русло новоутворені клітини крові і лімфоцити. Вважають, що клітини крові можуть також проникати у кровоносне русло крізь тимчасові пори в цитоплазмі ендотеліоцитів синусоїдних капілярів. Відростки макрофагів строми кісткового мозку мають здатність проникати між ендотеліоцитами у просвіт синусоїдних капілярів і фагоцитувати там чужорідні речовини. У подальшому В-лімфоцити заселяють В-залежні зони вторинних лімфоїдних органів. У зародка кровотворення починається з 19 доби у кров’яних острівцях жовткового мішка і продовжується у плода до початку 4-го місяця. У печінці кровотворення починається з 6-го тижня розвитку, а в селезінці – з 3-го місяця і триває до народження дитини.
Червоний кістковий мозок починає формуватися в кістках зародка в кінці 2-го місяця розвитку, а з 12-го тижня в кістковому мозку розвиваються кровоносні судини, зокрема, синусоїдні капіляри. Навколо судин розташовується ретикулярна тканина, формуються острівці кровотворення. З цього часу кістковий мозок починає функціонувати як орган кровотворення. Починаючи з 20-го тижня пренатального розвитку, маса кісткового мозку швидко збільшується, він проникає в епіфізи кісток і в утворену кістковомозкову порожнину.
У немовлят кістковомозкові порожнини всіх кісток заповнені червоним кістковим мозком. Перші жирові клітини утворюються в кістковому мозку через 1–6 місяців після народження дитини. Тільки з 4–5 років у кістковомозковій порожнині довгих кісток він поступово замінюється жовтим кістковим мозком. Жовтий кістковий мозок складається з численних жирових клітин – адипоцитів. Зрілі жирові клітини мають діаметр до 120 мкм і переважно кулясту форму. У таких клітинах вузький обідок цитоплазми і сплющене ядро відсунуті на периферію однією великою краплею жиру. У старечому віці жовтий кістковий мозок набуває драглистої консистенції і перетворюється на желатинозний кістковий мозок. У жовтому кістковому мозку кровотворення відсутнє. Однак при великих крововтратах у жовтому кістковому мозку можуть утворюватись острівці червоного кісткового мозку. У людей віком 20–25 років кістковомозкові порожнини довгих кісток заповнені тільки жовтим кістковим мозком. У дорослих людей в кістковомозкових порожнинах плоских кісток половину об’єму кісткового мозку займають жирові клітини.
Загруднинна залоза (тимус)
Загруднинна залоза, або тимус (thymus) є первинним лімфоїдним (імунним) органом, у якому відбувається антигеннезалежна проліферація і диференціація субпопуляцій Т-лімфоцитів із стовбурових клітин. У подальшому Т-лімфоцити з тимуса потрапляють у кров і заселяють тимусзалежні зони (Т-зони) вторинних лімфоїдних органів.
Тимус розташований за грудниною у верхньому середостінні між правою і лівою середостінними частинами пристінкової плеври – у верхньому міжплевральному полі. Він складається з двох асиметричних видовжених часток – правої та лівої часток (lobi dexter et sinister), які з’єднані між собою у середній частині. Нижня частина кожної частки розширена, досягає рівня IV ребрового хряща, а верхня частина часток звужена. Ліва частка в більшості випадків довша за праву. Переважно частки виступають в передню ділянку шиї у вигляді двозубої вилки. Ось чому раніше називали тимус вилочковою залозою.
Загруднинна залоза вкрита тонкою сполучнотканинною капсулою, від якої всередину органа відходять кіркові перегородки (septi corticales), що розділяють її на численні часточки загруднинної залози (lobuli thymi), розміри яких коливаються від 1 до 10 мм.
Численні епітеліоретикулоцити – особливі епітеліальні клітини зірчастої форми, з’єднуючись між собою довгими розгалуженими відростками, утворюють тривимірний каркас органа. В комірочках цієї сітки – каркаса містяться клітинні елементи паренхіми тимуса. Епітеліоретикулоцити мають велике кругле світле ядро, в якому містяться переважно еухроматин і чітке ядерце. Цитоплазми відносно мало, у ній поблизу ядра розташовано небагато пучків тонофіламентів, вони проникають у відростки епітеліоретикулоцитів і досягають десмосом, що утворюють міжклітинні контакти між відростками цих клітин. У цитоплазмі міститься помірна кількість мітохондрій, багато вільних рибосом і включень глікогену. У деяких ділянках цитоплазми добре розвинена гранулярна ендоплазматична сітка. Крім того, виявляються великі електроннощільні лізосомоподібні гарнули, а також дрібні щільні кулясті гранули, схожі на секреторні. У цитоплазмі присутні групи кулястих мембраних вакуоль, що містять аморфний матеріал помірної електронної щільності, який нагадує крапельки слизу келихоподібних екзокриноцитів. Окрім того, епітеліоретикулоцити синтезують біологічно активний поліпептид – тимозин (тимопоетин), що регулює диференціацію Т-лімфоцитів. Паренхіма загруднинної залози (тимуса) побудована в основному з різних субпопуляцій Т-лімфоцитів (тимоцитів), а також макрофагів, незначної кількості фібробластів, міофібробластів і тканинних базофілів. Периферійна частина часточки називається кірковою речовиною загруднинної залози (cortex thymi), а центральна ділянка часточки (на гістологічних препаратах дещо світліша) – мозковою речовиною загруднинної залози (medulla thymi).
У кірковій речовині компактно розміщені малі та середні лімфоцити в оточенні макрофагів. У кіркову речовину з червоного кісткового мозку з кров’ю потрапляють попередники Т-лімфоцитів, які під дією тимозину проліферують. У субкапсулярній зоні кіркової речовини є багато Т-лімфоцитів, зокрема, лімфобластів – великих клітин діаметром 12– 18 мкм. Лімфобласти мітотично поділяються і мігрують у мозкову речовину, поступово дозріваючи, перетворюючись у малі лімфоцити діаметром 6–7 мкм. Кожен лімфобласт у результаті шести послідовних мітотичних поділів утворює 128 малих лімфоцитів.
Мозкова речовина часточки тимуса утворена малими, середніми і великими Т-лімфоцитами, які оточені епітеліоретикулоцитами і макрофагами, але кількість цих клітин значно менша порівняно з кірковою речовиною. Характерною морфологічною ознакою загруднинної залози є наявність у мозковій речовині часточок особливих колових (концентричних) нашарувань епітеліоретикулярних клітин, які називаються тільцями тимуса (corpuscula thymi), або тільцями Гассаля, де замкнуті безсудинні кулясті структури діаметром 20–200 мкм. У тільцях тимуса сплощені епітеліоретикулярні клітини розміщені коловими шарами, нагадуючи цибулину. Клітини з’єднуються між собою численними десмосомами. У їх цитоплазмі містяться гранули кератогіаліну, кератину, пучки фібрил. Кератинізація клітин тілець посилюється в напрямку від їх периферії до центру. При нашаруванні на тільце тимуса нових клітин, живлення клітин, що містяться в центрі тільця, погіршується і вони дегенерують. Тому у центрі великих тілець Гассаля міститься клітинний детрит, оточений епітеліоретикулярними клітинами з великим вмістом кератину. Функція тілець ще невідома. У мозковій речовині часточок є багато посткапілярних венул, через стінки яких у кровоносне русло поступає рециркулюючий пул субпопуляцій Т-лімфоцитів.
Кровопостачання та іннервація тимуса
Тимус кровопостачають гілки загруднинної залози (rr. thymici), що відходять від внутрішньої грудної артерії, дуги аорти і плечо-головного стовбура. Ці артерії розгалужуються на міжчасточкові і внутрічасточкові судини, від яких відходять дугові артерії, що галузяться на капіляри. У кірковій речовині часточок капіляри формують численні дуги (аркади), які анастомозують між собою, після чого прямують у мозкову речовину, де переходять у посткапілярні венули мозкової речовини, що вистелені ендотеліоцитами призматичної форми (високим ендотелієм). У кірковій речовині часточок капіляри оточені тонким сполучнотканинним периваскулярним простором, що містить тонкі колагенові і ретикулярні волокна. Цей простір іззовні обмежований суцільним шаром епітеліоретикулоцитів, з’єднаних між собою десмосомами. У периваскулярному просторі містяться макрофаги, лімфоцити і циркулює міжклітинна рідина. Вени загруднинної залози (vv. thymici) впадають у плечо-головні вени і внутрішні грудні вени. Лімфатичні капіляри, які оточують часточки, формують лімфокапілярні сітки, а від них відходять лімфатичні судини, що впадають у трахео-бронхові і притрахейні лімфатичні вузли. Тимус іннервують гілки автономної частини периферійної нервової системи: парасимпатичні гілки блукаючих нервів і післявузлові симпатичні волокна, що відходять від шийно-грудного і верхнього грудного симпатичних вузлів.
Гематотимусний бар’єр
Однією з особливостей будови гемомікроциркуляторного русла тимуса є наявність у кірковій речовині часточок гематотимусного бар’єра. Він утворений стінкою гемокапілярів, периваскулярним простором і суцільним шаром розміщених на базальній мембрані епітеліоретикулоцитів, що супроводжує всі судини гемомікроциркуляторного русла, зокрема, посткапілярні венули. Такий бар’єр надійно захищає паренхіму тимуса від антигенів, що циркулюють у кровоносному руслі.
Розвиток і вікові особливості тимуса
Структуру і функцію тимуса неможливо зрозуміти, не знаючи його ембріонального розвитку. Загруднинна залоза починає формуватися у зародка на 4–5 тижні із епітеліальних клітин ІІІ і IV зябрових кишень з кожного боку, що у вигляді тяжів вростають каудально у мезенхіму. З епітеліальних клітин, що інтенсивно мітотично поділяються, утворюються епітеліоретикулоцити. На другому місяці ембріонального розвитку в епітеліальний зачаток тимуса вростають кровоносні капіляри, з яких у тканину проникають стовбурові клітини – попередники лімфоцитів. Ці клітини розташовані на периферії зачатка, інтенсивно мітотично поділяються, диференціюються у малі лімфоцити, а потім мігрують в глибину органа. Поділ тимуса на часточки, кіркову і мозкову речовини відбувається на третьому місяці розвитку. У немовлят маса загруднинної залози (тимуса) складає в середньому 13,3 г (від 7,7 г до 34,0 г). Упродовж перших трьох років життя дитини тимус росте найінтенсивніше. Загруднинна залоза досягає максимального розвитку у дітей і підлітків віком 12–16 років, тобто в період статевого дозрівання організму. Тоді її маса становить у середньому 37,5 г, а довжина 7,5–16 см. З віком маса тимуса поступово зменшується, у віці 16–20 років дорівнює в середньому 25,5 г, у людей віком 21–36 років – 22,3 г. У людей похилого віку маса тимуса становить 13–15 г.
Тимусні тільця (тільця Гассаля) діаметром 35– 40 мкм і більше вже наявні у загруднинній залозі немовлят – до 4–8 тілець на зрізі кожної часточки. Надалі їх кількість і розміри зростають. У 8-річних дітей діаметр тимусних тілець досягає 140–320 мкм. У дорослих людей дрібні тільця трапляються рідко.
У дітей і підлітків загруднинна залоза м’яка, сіро-рожевого кольору. До 10-річного віку в часточках тимуса переважає кіркова речовина, хоча починаючи з 3–4-річного віку товщина кіркової речовини зменшується, а її внутрішня межа стає нечіткою. У 10-річних дітей об’єм кіркової і мозкової речовин у часточках тимуса майже однаковий. Надалі поступово товщина кіркової речовини часточок зменшується, а об’єм мозкової речовини збільшується.
Одночасно з перебудовою і зміною співвідношення кіркової та мозкової речовин у паренхімі тимуса утворюється жирова тканина, а також розростається сполучна тканина. Якщо у немовлят сполучна і жирова тканини складають тільки 7 % від маси тимуса, то у 20-річних людей вона досягає вже 40 % і надалі її кількість зростає. У людей похилого віку лімфоїдна тканина тимуса повністю не зникає, вона представлена окремими острівцями, які оточені жировою і сполучною тканинами, які складають до 90 % маси органа.
Відсутність вікової інволюції загруднинної залози призводить до важкого захворювання – тиміко-лімфатичного статусу. При цьому зменшується опірність організму до інфекцій та інтоксикацій, зростає загроза виникнення злоякісних утворів. При дії на організм різноманітних стресів, при травмах, інтоксикаціях, інфекціях тощо може настати так звана акцидентальна інволюція тимуса. При цьому за рахунок апоптозу масово гинуть лімфоцити (переважно кіркової речовини), частина Т-лімфоцитів швидко надходить у кров і мігрує у вторинні лімфоїдні органи, проліферують епітеліоретикулоцити, внаслідок чого зникає межа між кірковою та мозковою речовиною у часточках. Такі морфологічні зміни є проявом захисних реакцій організму.
ВТОРИННІ ЛІМФОЇДНІ (ІМУННІ) ОРГАНИ
Лімфатичні вузли
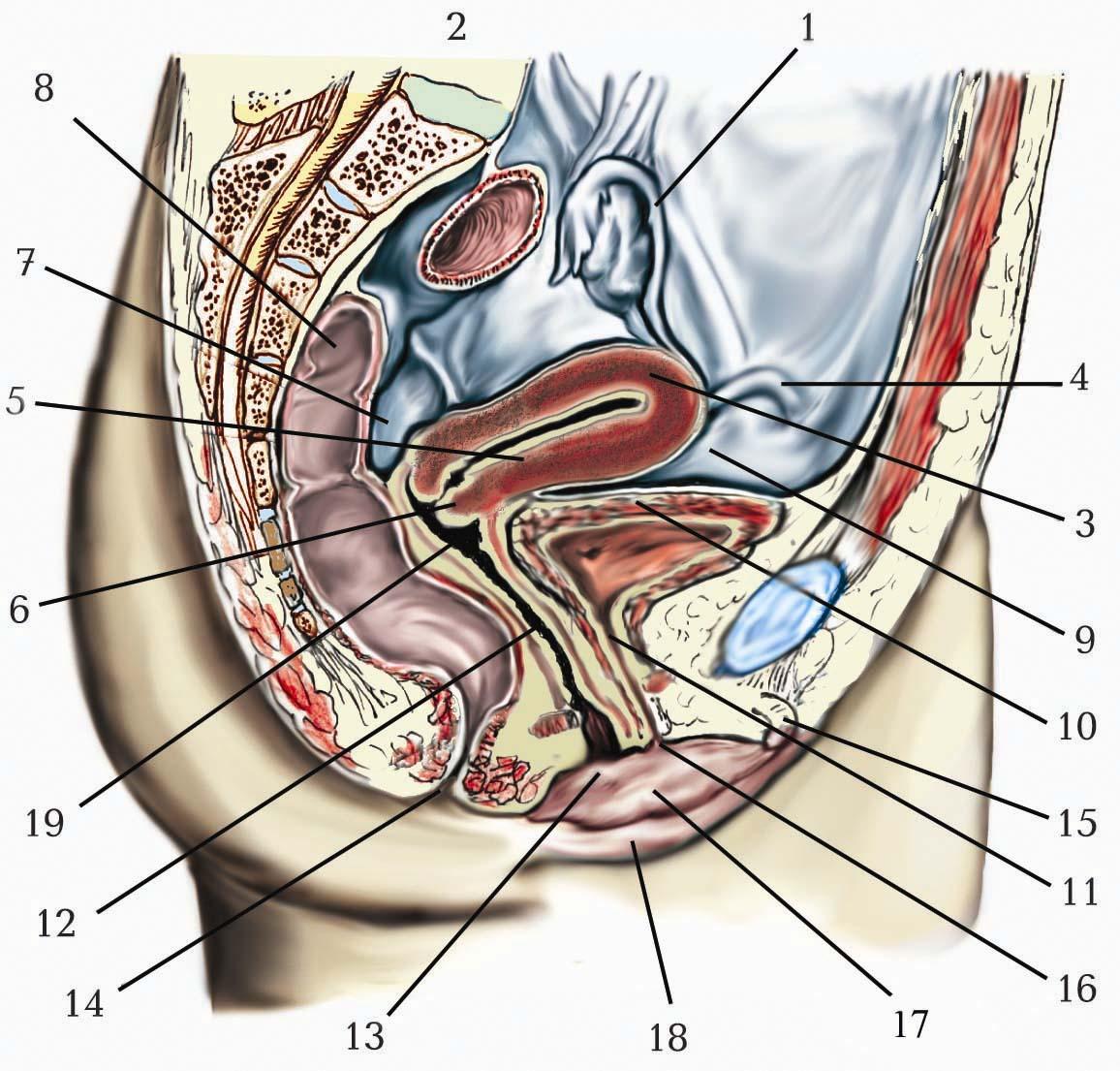
Лімфатичні вузли (nodi lymphаtici; nodi lymphoidei; lymphonodi) є вторинними лімфоїдними (імунними) органами, що розташовані на шляхах протікання лімфи від органів і тканин організму до лімфатичних стовбурів і проток . Лімфатичні вузли є біологічними „фільтрами”, в яких знешкоджуються антигени, відбувається антигензалежна проліферація і диференціація Ті В-лімфоцитів і формується конкретна імунна відповідь на їхню дію.
Лімфатичні вузли мають різноманітну форму: кулясту, овоїдну, стрічкоподібну, але найчастіше – бобоподібну. Їх розміри коливаються від 0,5 мм до 50 мм і більше. Лімфатичні вузли розташовані в пухкій сполучній тканині, найчастіше групами, у яких налічується до кількох десятків вузлів. Вузли, які приймають лімфу від певної ділянки організму, називаються ділянковими лімфатичними вузлами. Вони поділяються на пристінкові, нутрощеві та змішані. Відповідно у пристінкові лімфатичні вузли впадають приносні лімфатичні судини від шкіри, підшкірної клітковини і опорно-рухового апарату, в нутрощеві – від внутрішніх органів, а в змішані вузли – від тіла і внутрішніх органів.
Лімфатичний вузол (nodus lymphаticus; nodus lymphoideus; lymphonodus) вкритий сполучнотканинною капсулою лімфатичного вузла (capsula lymphonodi), від якої вглиб органа відходять перекладки, або трабекули лімфатичного вузла (trabeculae lymphonodi), вони бувають кірковими і мозковими. Перекладки мають різноманітну форму і утворюють своєрідний сполучнотканинний каркас. У перекладках проходять судини і нерви. Простори між перекладками заповнені стромою, що утворена ретикулярними клітинами і ретикулярними волокнами. У такій тривимірній сітці зі строми розташована лімфоїдна паренхіма, що складається з клітин лімфоїдного ряду, макрофагів тощо.
У найопуклішу частину лімфатичного вузла впадає 4–8, а інколи й більше приносних лімфатичних судин (vasa lymphаtica afferentia), які відкриваються у крайову пазуху, або підкапсульну пазуху (sinus marginalis; sinus subcapsularis). З протилежного боку вузол має невелике втиснення – ворота лімфатичного вузла (hilum lymphonodi), через які проходять кровоносні судини і нерви, а також виходять 2–4 виносні лімфатичні судини (vasa lymphatica efferentia), що прямують до наступного лімфатичного вузла або лімфатичного колектора.
Паренхіма лімфатичного вузла складається з кіркової речовини лімфатичного вузла (cortex lymphonodi), яка розташована ближче до капсули і займає периферійні відділи вузла, і з мозкової речовини лімфатичного вузла (medulla lymphonodi), що розташована в центральній частині вузла ближче до його воріт. Паренхіма вузла складається із структурних компонентів, що належать до Ті В-залежних зон.
Паренхіма лімфатичного вузла пронизана численними вузькими каналами – лімфатичними проміжними пазухами (sinus intermedii lymphatici), по яких лімфа протікає від крайової пазухи до ворітної пазухи. У пазухи з паренхіми вузла потрапляють субпопуляції Ті В-лімфоцитів. Усі пазухи вистелені плоскими ендотеліальними клітинами. Через кіркову речовину проходять кіркові проміжні лімфатичні пазухи (sinus intermedii lymphatici corticales), які з одного боку межують з кірковою перекладкою, а з іншого – оточені паренхімою. У мозковій речови-ні розташовані мозкові проміжні лімфатичні пазухи (sinus intermediі lymphatici medullares) різноманітної форми. Одні мозкові проміжні пазухи розміщені у мозковій паренхімі – мозкових тяжах. Інші мозкові проміжні пазухи оточують з усіх боків мозкову перекладку, а ззовні від них розташовані мозкові тяжі.
Між ендотеліоцитами, що вистеляють лімфатичні пазухи, можуть проникати з паренхіми лімфатичного вузла в лімфу і навпаки лімфоцити, макрофаги та інші клітини. У просвіті пазух міститься дрібнопетлиста сітка, утворена з ретикулярних волокон і клітин. У петлях цієї сітки можуть затримуватись сторонні частинки (вугільний і тютюновий пил), мікроби, злоякісні клітини тощо, які потрапили в лімфатичний вузол разом з лімфою. Часточки пилу переносяться макрофагами в паренхіму лімфатичного вузла і там відкладаються. Залишки зруйнованих клітин, що потрапили в лімфу, знищуються, а із злоякісних клітин можуть виникати вторинні пухлини – метастази.
У периферійній частині кори розташовані численні лімфоїдні вузлики (noduli lymphoidei) діамером 0,5–1 мм – це скупчення лімфоїдних клітин, в основному В-лімфоцитів, тому ці структури називають В-залежними зонами. Розрізняють первинні лімфоїдні вузлики (noduli lymphoidei primarii), які не мають зародкового центру, або центру розмноження (centrum germinativum), бо щільність лімфоцитів у них однакова і вторинних лімфоїдних вузликів (noduli lymphoidei secundarii) із зародковим центром, або центром розмноження (centrum germinativum). У центрах розмноження відбувається антигензалежна проліферація і диференціація різноманітних субпопуляцій Ті В-лімфоцитів. Там є багато лімфобластів і клітин, що мітотично поділяються, а також середніх лімфоцитів, тому на гістологічних препаратах ці центри видаються світлішими. Периферійна частина лімфоїдних вузликів називається крайовою зоною, або плащовою зоною (zona marginalis). На гістологічних зрізах крайова зона забарвлена інтенсивніше (темніше), бо щільно заповнена малими і середніми лімфоцитами. Через крайову зону постійно мігрують лімфоїдні клітини, макрофаги та інші клітини. Зокрема, “молоді” плазматичні клітини виходять із центра розмноження і мігрують через прикіркову ділянку (паракортикальну зону), у мозкові тяжі, де вони контактують з іншими лімфоцитами, макрофагами і ретикулярними клітинами, обмінюючись з ними відповідною інформацією. Кількість вторинних лімфоїдних вузликів із центром розмноження збільшується після дії антигенів.
Частина периферійної кори, що розміщена між лімфоїдними вузликами і крайовою пазухою, називається підкапсульною частиною, або крайовою частиною (pars subcapsularis; pars marginalis). Дифузна лімфоїдна тканина, яка розміщена між лімфоїдними вузликами, утворює міжвузликову частину (pars internodularis). Тут розташовані в основному малі і середні лімфоцити. Глибокі відділи кіркової речовини, що межують з мозковою речовиною, називаються прикірковою ділянкою (paracortex), раніше її називали паракортикальною зоною. Це Т-залежна зона, бо там містяться переважно малі субпопуляції Т-лімфоцитів. Характерною морфологічною особливістю цієї зони є наявність в ній численних венул з високим ендотелієм (venulae altoendotheliales), через які здійснюється рециркуляція лімфоцитів у паренхіму лімфатичного вузла із крові.
Паренхіма мозкової речовини представлена мозковими тяжами (chordae medullares), які мають різноманітну форму і напрямок, а на гістологічному зрізі утворюють складні переплетення і острівці, що анастомозують між собою. У мозкових тяжах переважають В-лімфоцити. Зокрема, там розташовані “зрілі” плазмоцити (В-ефектори), які виконують свою основну функцію – синтезують антитіла, обов’язковою є також присутність макрофагів, тому мозкові тяжі належать до В-залежної зони.
У структурних компонентах лімфатичних вузлів постійно відбуваються процеси міграції, диференціації та проліферації субпопуляцій Ті В-лімфоцитів. Значна частина лімфоцитів з лімфою через виносні лімфатичні судини потрапляє у наступний лімфатичний вузол або у лімфатичні стовбури і протоки, що впадають у венозні судини. У подальшому лімфоцити рециркулюють із крові у вторинні лімфоїдні органи і тканини.
Розвиток і вікові особливості лімфатичних вузлів
Лімфатичні вузли починають утворюватися у зародка на 5–6 тижні розвитку з мезенхіми, що оточує новоутворені кровоносні і лімфатичні судини. Meзенхіма проникає у просвіт лімфатичної судини, а її клітини диференціюються в паренхіму вузла. Просвіт лімфатичної судини перетворюється на крайову пазуху, а з її розгалужень виникають проміжні пазухи. Починаючи з 19-го тижня розвитку, помітна межа між кірковою і мозковою речовинами. Дещо пізніше утворюються лімфоїдні вузлики. Центри розмноження виникають у вузликах перед народженням дитини. Утворення лімфатичних вузлів у різних ділянках тіла людини відбувається впродовж усього пренатального періоду розвитку організму і навіть після народження. Основні формоутворюючі процеси в лімфатичних вузлах завершуються у дітей 10–12 років. Інволютивні зміни (зменшення кількості лімфоїдної тканини, збільшення сполучної і жирової тканин) починаються в лімфатичних вузлах вже в юнацькому віці. З віком зменшується кількість і розміри лімфатичних вузлів. Вони можуть зростатися між собою, утворюючи ланцюжки або сегментований крупніший вузол.
Кровопостачання та іннервація лімфатичних вузлів
Кожен лімфатичний вузол кровопостачають від 1–2 до 10 дрібних гілочок, що відходять від найближчих артерій. Зайшовши в лімфатичний вузол через його ворота, артерії проходять у складі його сполучнотканинних перекладок, галузяться і заходять у кожну структурно-функціональну зону паренхіми. Там вони розгалужуються до капілярів, які утворюють сіткоподібні структури і збираються у закапілярні венули. Венули, зливаючись, утворюють вени, які супроводжують артерії, виходять з лімфатичного вузла через його ворота і впадають у найближчі вени. Ще раз наголосимо, що найбільше венул з високим ендотелієм міститься у прикірковій ділянці вузла, через які в основному відбувається рециркуляція лімфоцитів з крові у паренхіму лімфатичного вузла.
Лімфатичні вузли, як і кровоносні судини, іннервують завузлові волокна симпатичної частини автономного відділу периферійної нервової системи, що відходять від відповідних симпатичних вузлів. Симпатичні завузлові волокна, обплітаючи артерії, заходять з ними в лімфатичний вузол, де іннервують усі його структурні компоненти. Чутлива інформація від лімфатичних вузлів йде по чутливих волокнах відповідних нервів.
Селезінка
Селезінка (lien, splen) є вторинним лімфоїдним (імунним) органом, який забезпечує імунний контроль крові, що протікає від аорти в систему ворітної печінкової вени. У селезінці знищуються „відпрацьовані” еритроцити, клітини крові, антигени та інші чужорідні часточки. У ній відбувається антигензалежна проліферація і диференціація популяцій Ті В-лімфоцитів. Селезінка ) розташована у черевній порожнині – в лівому підребер’ї, у верхньому поверсі очеревинної порожнини в селезінковому закутку на рівні ІХ–ХІ ребер. Поздовжня вісь селезінки відповідає нахилу цих ребер.
Маса селезінки дорослої людини становить 150– 200 г, вона має видовжену і опуклу форму (тому її за подібністю в народі називають косою), її довжина дорівнює 10–14 см, ширина – 6–10 см і товщина – 3–4 см. Селезінка темно-червоного кольору і м’яка на дотик.
У селезінці виділяють дві поверхні: гладку і опуклу діафрагмову поверхню (facies diaphragmatica), що обернена вбік і вверх до діафрагми, і нерівну передньоприсередню нутрощеву поверхню (facies visceralis). На нутрощевій поверхні розташовані видовжені селезінкові ворота (hilum splenicum; hilum lienale), через які проходять судини і нерви. Нутрощева поверхня має окремі ділянки, до яких дотикаються сусідні внутрішні органи: шлункова поверхня (facies gastrica), до якої прилягає дно шлунка; ниркова поверхня (facies renalis), що розміщується позаду селезінкових воріт, до неї прилягає верхній кінець (полюс) лівої нирки і ліва надниркова залоза; ободовокишкова поверхня (facies colica), що розташована нижче від селезінкових воріт і ближче до її переднього кінця, вона стикається з лівим згином ободової кишки. Дещо вище від ободовокишкової поверхні, відразу за селезінковими воротами, до селезінки прилягає хвіст підшлункової залози. Верхній край (margo superior) селезінки гострий, спрямований допереду і відокремлює її шлункову поверхню від діафрагмової. На цьому краї помітні 2–3 неглибокі поперечні виїмки. Нижній край (margo inferior) селезінки дещо заокруглений. Обидва краї сходяться на передньому і задньому кінцях селезінки. Передній кінець (extremitas anterior) гостріший, спрямований донизу і вперед до лівої ребрової дуги, міститься дещо вище поперечної ободової кишки. Заокруглений задній кінець (extremitas posterior) направлений догори і назад до хребта, не досягаючи до нього на 4–5 см.
Селезінка вкрита нутрощевою очеревиною з усіх боків (інтраперитонеально), за винятком її воріт. Очеревина утворює зв’язки, що фіксують селезінку в ділянці її воріт і прикріплюються до відповідних органів: шлунково-селезінкову зв’язку (lig. gastrospleni-
Очеревина міцно зростається з з капсулою, або
волокнистою оболонкою (capsula; tunica fibrosa) селезінки, (capsula splenica; tunica fibrosa splenica), від якої всередину органа відходять селезінкові перекладки (trabeculae splenicae). Сполучна тканина капсули і селезінкових перекладок містить, окрім пучків колагенових та еластичних волокон і фібробластів, небагато гладких міоцитів. Гладкі міоцити строми, скорочуючись, сприяють зменшенню об’єму селезінки і викиду депонованої крові у селезінкову вену. Окрім перекладок, сполучнотканинний каркас селезінки утворює її строма з ретикулярних волокон і ретикулярних клітин. У комірках цього каркасу розташована паренхіма – селезінкова пульпа (pulpa splenica; pulpa lienalis). Розрізняють червону пульпу і білу пульпу.
Червона пульпа (pulpa rubra) переважає, її маса становить приблизно 75–80 % від маси селезінки. Вона представлена скупченнями клітин крові, що оточені ретикулярними клітинами або містяться в системі венозних пазух селезінки. Ділянки червоної пульпи, що розташовані між венозними пазухами, називаються селезінковими тяжами (chordae lienis). У тяжах В-лімфоцити перетворюються у плазмоцити, а моноцити – у макрофаги. Макрофаги селезінки впізнають і руйнують старі або пошкоджені еритроцити та тромбоцити.
Біла пульпа (pulpa alba) становить 20–25 % від маси селезінки і у вигляді острівців розташована серед червоної пульпи. Біла пульпа – це лімфоїдна тканина, що утворена з лімфоцитів, плазмоцитів, макрофагів, дендритних та інтердигітуючих клітин. Біла пульпа виконує функцію вторинного лімфоїдного органа. Виділяють такі структурні компоненти білої пульпи: лімфоїдні вузлики селезінки і навколоартеріальні лімфоїдні піхви.
Лімфоїдні вузлики селезінки (noduli lymphoidei splenici) мають діаметр 0,3–0,5 мм і складаються з чотирьох зон: навколоартеріальної, мантійної, крайової і зародкового центру, або центру розмноження. У центрах розмноження лімфоїдних вузликів селезінки, як і в лімфатичному вузлі, є багато В-лімфобластів, клітин, що мітотично поділяються, макрофагів, плазмоцитів. Там відбувається антигензалежна проліферація і диференціація субпопуляцій лімфоцитів. Навколоартеріальна зона представлена скупченням Т-лімфоцитів і макрофагів – інтердигітуючими антигенпрезентуючими клітинами навколо центральної артерії, що пронизує лімфоїдний вузлик ексцентрично. Ця ділянка є аналогом прикіркової ділянки лімфатичних вузлів (Т-зона). Темніша мантійна зона, що оточує центр розмноження лімфоїдного вузлика, утворена із щільно розміщених малих В-лімфоцитів, незначної кількості Т-лімфоцитів, плазмоцитів і макрофагів. Крайова зона лімфоїдного вузлика розташована на межі з червоною пульпою і оточена синусоїдними гемокапілярами. У цій зоні розташовані Ті В-лімфоцити і макрофаги. Після дозрівання лімфоцити мігрують із центру розмноження в мантійну і крайову зони, а потім потрапляють у кровоносне русло.
Навколоартеріальні лімфоїдні піхви (vaginae lymphoidei periarteriales) – це видовжені скупчення лімфоцитів, які, як муфти, охоплюють артерії білої пульпи селезінки, пeрeходячи у лімфоїдні вузлики. У центральній частині піхви, що прилягає до артерії, розташовані в основному В-лімфоцити і плазмоцити, на периферії – Т-лімфоцити.
Кровопостачання та іннервація селезінки
Для того щоб зрозуміти будову і функцію селезінки, необхідно знати систему її кровопостачання.
Селезінка має особливу систему кровообігу, що забезпечує її функцію. Селезінкова артерія (a. splenica; a. lienalis) ще перед входженням у селезінку розгалужується на декілька гілок, що входять в її ворота. Ці гілки галузяться на 4–5 сегментних артерій (aa. segmentales), а вони у свою чергу – на перекладкові артерії (aa. trabeculares), що проходять у перекладках селезінки, а потім входять в її паренхіму. У перекладкових артеріях добре розвинена середня м’язова оболонка, особливо коловий (спіральний) шар гладких міоцитів. Артерії, що галузяться в селезінковій пульпі, називаються пульпарними артеріями (aa. pulpares) і мають діаметр до 0,2 мм. Навколо них у білій пульпі розташовані навколоартеріальні лімфоїдні піхви. Потім ці артерії проходять через лімфоїдні вузлики і вже називаються центральними артеріями (aa. centrales), їх діаметр дорівнює приблизно 50 мкм. Внутрішня еластична мембрана відсутня в їхній стінці, а ендотелій стоншується. Вийшовши з лімфоїдного вузлика, центральна артерія розгалужується в червоній пульпі на 2–6 китичкових (пензликових) артеріол (arteriolae penicillares), або китиць (penicilli) діаметром до 50 мкм. Китичкові артеріоли оточені еліпсоподібними муфтами – скупченням макрофагів, лімфоцитів і ретикулярних клітин. Тому такі муфти називають навколоартеріолярними макрофагоцитарними піхвами (vaginae periarteriolares macrophagоcyticae), а артеріоли, що проходять через них, – еліпсоїдними артеріолами. Ретикулярні клітини і волокна цих структур є своєрідними артеріальними замикачами (сфінктерами) селезінки. За будовою стінки ці артеріоли подібні до гемокапілярів. Китичкові артеріоли розгалужуються на пористі капіляри (vasa capillaria) діаметром 4–6 мкм, які частково впадають у венозні пазухи (синусоїди) селезінки (sinus venulares lienis), або синусоїдні судини селезінки (vasa sinusoidea lienis). Більша частина капілярів відкривається безпосередньо у червону пульпу, формуючи систему відкритого кровообігу селезінки. Ендотеліоцити венозних пазух селезінки мають вигляд паличкоподібних клітин, з’єднаних своїми бічними відростками, щілиноподібний простір між якими забезпечує можливість міграції клітин крові, що вільно вийшли з капілярів у червону пульпу, мігрують зворотно через вузькі міжендотеліальні щілини у венозні пазухи. Нормальні еритроцити, завдяки гнучкості елементів цитоскелета, легко деформуються і без затримки потрапляють у просвіт пазух. Старі або пошкоджені еритроцити не деформуються, залишаються у червоній пульпі і в подальшому підлягають фагоцитозу. Венозні пазухи виконують також функцію депо крові. При переході венозних пазух у вени червоної пульпи колові (циркулярні) скупчення гладких міоцитів формують венозні замикачі (сфінктери) селезінки. Отже, такі замикачі регулюють кровообіг у селезінці. Вени червоної пульпи впадають у перекладкові вени (vv. trabeculares), що формують селезінкову вену (v. splenica; v. lienalis), по якій відтікає венозна кров у ворітну печінкову вену (v. portae hepatis).
Розвиток і вікові особливості селезінки
Закладається селезінка у зародка на 5–6-му тижні розвитку у вигляді скупчення клітин мезенхіми в дорсальній брижі. Згодом у цьому мезенхімному скупченні утворюються щілини – майбутні судини селезінки, навколо яких диференціюються клітини пульпи. На 2–4-му місяці розвитку формуються венозні пазухи та інші ланки кровоносних судин. У цей час від капсули всередину органа вростають тяжі, що утворюють перекладки. Навколо венозних пазух виникають острівці кровотворення (гемопоезу). На 3–5-му місяці утворюються усі структурні елементи білої пульпи селезінки, зокрема, в лімфоїдних вузликах виникають центри розмноження. На 6-му місяці вже можна розрізнити червону пульпу. Із 6-го місяця інтенсивність мієлоїдного кровотворення зменшується, перед народженням дитини ця функція селезінки припиняється. Але інтенсивність лімфоцитопоезу наростає. У немовлят маса селезінки дорівнює в середньому 9,5 г, а біла пульпа складає 5–10 % від маси органа. У 3-місячних дітей маса селезінки зростає до 11–14 г, в однорічних вона становить 24–28 г, а обсяг білої пульпи в цей період є найбільшим – до 20,9 %. У 6-річних дітей маса селезінки становить 48–56 г, у 10-річних досягає 66–70 г, а в 16–17-річних підлітків дорівнює 165–171 г. Упродовж життя відносна кількість червоної пульпи не змінюється, коливається в межах 82–85 %. Кількість білої пульпи з віком зменшується. Якщо у 10-річних дітей біла пульпа займає в середньому 18,6 %, а в людей віком 21–30 років її вміст зменшується до 7,7–9,6 %, то в 50-річних людей її кількість не перевищує 6,5 % від маси селезінки.
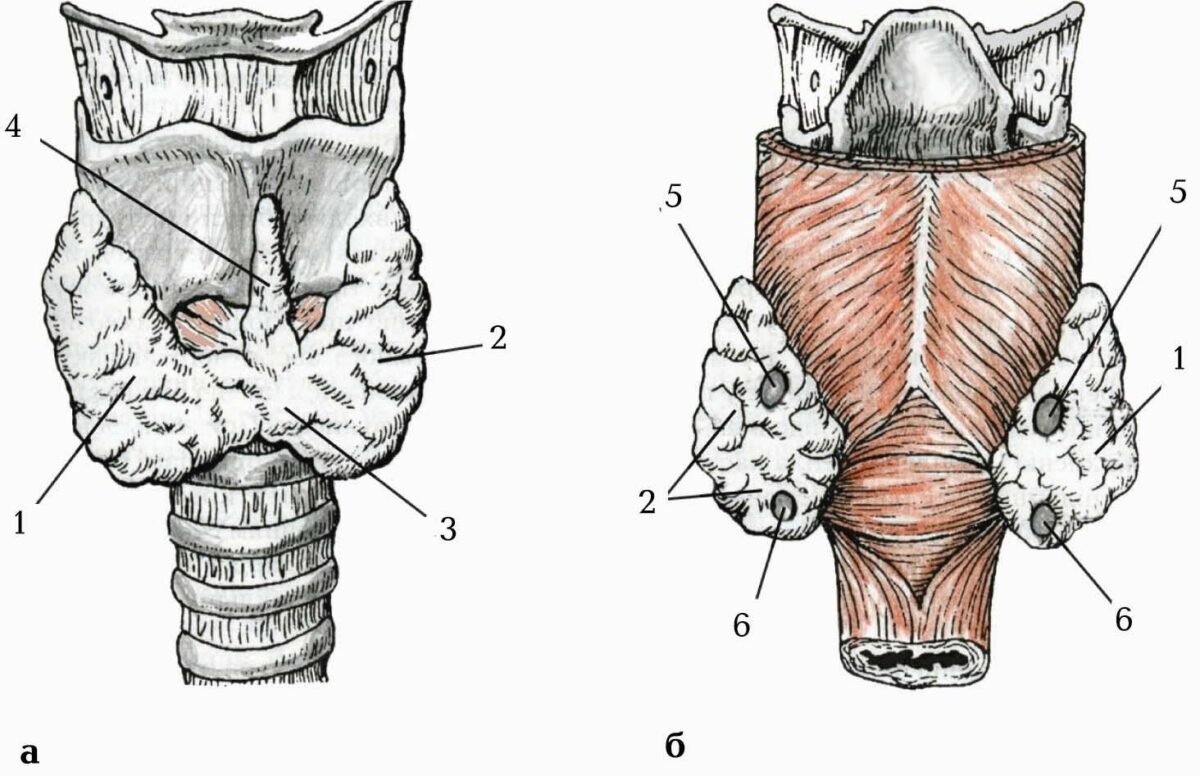
Мигдалики
Мигдалики (tonsillae) – це вторинні лімфоїдні органи, які розташовані в ділянці зіва, кореня язика і носової частини глотки. Є шість мигдаликів: два парні – піднебінний і трубний, та два непарні – язиковий і глотковий. Утворюючи лімфатичне (лімфоїдне) кільце глотки (anulus lymphoideus pharyngis) – кільце Пирогова – Вальдейєра, мигдалики оточують вхід у глотку з носової та ротової порожнин, куди з повітрям та їжею потрапляють і антигени. Структурно-функціональною одиницею мигдаликів є лімфоїдні вузлики (noduli lymphoidei), які за будовою та функцією подібні до лімфоїдних вузликів лімфатичного вузла.
Язиковий мигдалик (tonsilla lingualis) розташований у власній пластинці слизової оболонки кореня язика і утворений численними (80–90) лімфоїдними вузликами діаметром 1–4 мм. Мигдалик має овальну форму з максимальною довжиною до 25 мм у 14– 20-річному віці людини. Слизова оболонка спинки язика над мигдаликом нерівна, вона має від 60 до 150 підвищень – горбків, діаметр яких дорівнює 3–4 мм. Між горбками у слизовій оболонці є заглибини – мигдаликові крипти (cryptae tonsillares) глибиною 2–4 мм. Стінка крипт побудована з багатошарового плоского незроговілого епітелію, що інфільтрований лімфоцитами. У мигдаликові крипти відкриваються також протоки малих слинних залоз.
Язиковий мигдалик утворюється у плода на 6–7-му місяці пренатального розвитку, на 8–9-му місяці диференціюються лімфоїдні вузлики. Центри розмноження у вузликах утворюються тільки після народження дитини впродовж першого місяця життя. З віком кількість лімфоїдних вузликів зростає: у грудному віці – до 66, у періоді першого дитинства – до 85, у підлітків – до 90. Діаметр вузликів у цей період дорівнює 2–4 мм. Починаючи з юнацького віку, кількість лімфоїдних вузликів поступово меншає.
Кровопостачання та іннервація язикового мигдалика. Язиковий мигдалик живлять гілки правої і лівої язикових артерій, а венозна кров від нього відтікає в язикові вени. Лімфа від язикового мигдалика відтікає в ділянкові лімфатичні вузли – бічні глибокі шийні вузли.
Іннервують язиковий мигдалик парасимпатичні гілки язико-глоткових і блукаючих нервів (ІХ–Х пари черепних нервів), а чутливу інформацію від мигдалика передають чутливі волокна цих нервів та симпатичні завузлові волокна правого і лівого зовнішніх сонних сплетень, що проникають у мигдалик з гілками язикових артерій.
Піднебінний мигдалик (tonsilla palatina) – парний, видовженої овоїдної форми, розташований у мигдаликовій ямці м’якого піднебіння, що обмежована піднебінно-язиковою і піднебінно-глотковою дужками. Бічна поверхня мигдалика прилягає до сполучнотканинної підслизової основи стінки глотки, яку в цьому місці називають мигдаликовою капсулою (capsula tonsillaris). Від неї відходять перекладки вглиб мигдалика, що розділяють його на часточки. Приблизно на відстані 1 см збоку від піднебінного мигдалика проходить внутрішня сонна артерія. Присередня поверхня мигдалика вкрита багатошаровим плоским незроговілим епітелієм, на цій поверхні є до 20 мигдаликових ямочок (fossulae tonsillares), у які відкриваються мигдаликові крипти (cryptae tonsillares). Паренхіма мигдалика утворена численними лімфоїдними вузликами (noduli lymphoidei) діаметром 0,1–1,2 мм, у найбільших з них є центри розмноження. Між вузликами розташована лімфоїдна тканина у вигляді тяжів товщиною до 1–2 мм. Слизова оболонка над вузликами випинається та інфільтрована лімфоцитами.
Піднебінні мигдалики закладаються у плода на 12–14 тижні розвитку у вигляді скупчення мезенхіми під епітелієм другої глоткової кишені. На 5-му місяці в лімфоїдну тканину вростають епітеліальні тяжі, з яких згодом формуються крипти. Кількість лімфоїдної тканини у мигдалику більшає, перед народженням дитини утворюються одинокі лімфоїдні вузлики, але центри розмноження в них виникають вже після народження. Упродовж першого року життя дитини розміри мигдалика подвоюються – до 15 мм завдовжки і 12 мм завширшки. Найбільших розмірів піднебінний мигдалик досягає у дітей 8–13 років, тоді він має довжину до 28 мм, а ширину до 22 мм. Після 25–) 30 років у людей наступає вікова інволюція лімфоїдної тканини, а кількість сполучної тканини зростає.
Кровопостачання та іннервація піднебінних мигдаликів. Піднебінний мигдалик живлять мигдаликові гілки (rr. tonsillares) висхідної піднебінної артерії (гілка лицевої артерії), висхідної глоткової артерії (гілка зовнішньої сонної артерії), а також низхідної піднебінної артерії (гілка верхньощелепної артерії). Венозна кров від мигдалика відтікає по 3–4 мигдаликових венах (vv. tonsillares) у крилоподібне сплетення, а з нього – у занижньощелепну вену. Лімфа від внутрішньоорганних лімфокапілярних сіток, які оточують лімфоїдні вузлики, по кількох лімфатичних судинах, що прямують вбік, відтікає у ділянкові лімфатичні вузли – у бічні глибокі шийні вузли.
Іннервація піднебінних мигдаликів: чутлива інформація від кожного мигдалика передається по мигдаликових гілках (rr. Tonsillares язико-глоткового нерва (ІХ черепний нерв) та піднебінного нерва, що є гілками верхньощелепного нерва (гілка трійчастого нерва; V черепний нерв); парасимпатичні завузлові волокна, які відходять від крило-піднебінного вузла, що є складовими великого кам’янистого нерва (гілка лицевого нерва; VII черепний нерв); симпатичні завузлові волокна, які відходять від внутрішнього сонного сплетення і проникають у піднебінний мигдалик разом з артеріальними гілками.
Глотковий мигдалик (tonsilla pharyngea), або аденоїдний мигдалик (tonsilla adenoidea), розташований у власній пластинці слизової оболонки носової частини глотки у місці переходу її склепіння у задню стінку між глотковими отворами правої і лівої слухових труб. Тут слизова оболонка утворює 4–6 поперечних і косих складок, у товщі яких міститься паренхіма глоткового мигдалика, що складається з дифузної лімфоїдної тканини і лімфоїдних вузликів (noduli lymphoidei) діаметром до 0,8 мм. Між складками є відносно глибокі борозни, які відкриті донизу, в їхній просвіт відкриваються глоткові залози. У ділянці склепіння глотки складки низькі, між ними по серединній лінії проходить неглибока борозна. Слизoва оболонка над глотковим мигдаликом вкрита багатошаровим плоским незроговілим епітелієм. На його поверхні є мигдаликові ямочки (fossulae tonsillares), що переходять у мигдаликові крипти (cryptae tonsillares). Слизова оболонка в ділянках борозен і мигдаликових крипт вкрита одношаровим багаторядним війчастим епітелієм респіраторного типу, як і в інших відділах носової частини глотки. Сполучнотканинна строма глоткового мигдалика зростається з глотково-основною фасцією, що є похідною підслизового прошарку носової частини глотки.
Інколи у дітей і підлітків при запаленні глоткового мигдалика (аденоїдит) або при його збільшенні – гіпертрофії (аденоїди) складки слизової оболонки у цій ділянці дуже великі і звисають вниз, прикриваючи хоани. Це призводить до затрудненого носового дихання, а згодом і до виникнення аденоїдизму – синдрому, який є наслідком значного збільшення глоткового мигдалика.
Глотковий мигдалик починає утворюватися у плода на 3–4-му місяці розвитку у відповідній ділянці слизової оболонки. У немовлят глотковий мигдалик уже добре виражений, його довжина дорівнює 5–7 мм, а ширина – 5–6 мм. Упродовж першого року життя мигдалик росте дуже інтенсивно, у ньому формуються лімфоїдні вузлики. У кінці року довжина мигдалика становить 12 мм, а ширина – 6–10 мм. Найбільших розмірів глотковий мигдалик досягає у людини у віці 8–20 років: довжина – 13–21 мм, ширина 10–15 мм, а згодом наступає вікова інволюція.
Кровопостачання та іннервація глоткового мигдалика. Глотковий мигдалик живлять мигдаликові гілки (rr. tonsillares) правої і лівої висхідних глоткових артерій, що відходять від зовнішніх сонних артерій. Венозна кров від мигдалика відтікає у глоткове сплетення, а з нього – в обидві внутрішні яремні вени. Лімфа відтікає від глоткового мигдалика в заглоткові лімфатичні вузли, що входять до складу передніх глибоких шийних лімфатичних вузлів.
Іннервують глотковий мигдалик: чутлива інформація від мигдалика передається по мигдаликових гілках (rr. tonsillares) язико-глоткових нервів (ІХ пара черепних нервів), а також блукаючих нервів (Х пара черепних нервів); парасимпатичні завузлові волокна відходять від крило-піднебінного вузла (VII пара черепних нервів); симпатичні завузлові волокна відходять від зовнішнього сонного сплетення і проникають у глотковий мигдалик разом з артеріальними гілками.
Трубний мигдалик (tonsilla tubaria) – парний, розташований у власній пластинці слизової оболонки носової частини глотки в ділянці трубного валка, що обмежовує позаду глотковий отвір слухової труби. Слизова оболонка над мигдаликом вкрита одношаровим багаторядним війчастим епітелієм респіраторного типу; у ній є мигдаликові крипти (cryptae tonsillares). Паренхіма мигдалика складається з дифузної лімфоїдної тканини та лімфоїдних вузликів (noduli lymphoidei), слизова оболонка над ними утворює невеликі горбки.
Трубний мигдалик починає розвиватися у плода на 7–8-му місяці у товщі слизової оболонки навколо глоткового отвору слухової труби, в якій диференціюється лімфоїдна тканина, а лімфоїдні вузлики і центри розмноження утворюються впродовж першого року життя дитини. У немовлят довжина мигдалика становить 7,5 мм, а найбільшого розвитку мигдалик досягає у дітей віком 4–7 років, тоді його довжина становить 15–20 мм. Вікова інволюція трубних мигдаликів починається у підлітковому віці.
Кровопостачання та іннервація трубних мигдаликів. Трубний мигдалик кровопостачають мигдаликові гілки (rr. tonsillares) висхідної глоткової артерії, що відходить від зовнішньої сонної артерії, а венозна кров від мигдалика відтікає у глоткове сплетення, а з нього – у внутрішню яремну вену. Лімфа відтікає від трубного мигдалика в заглоткові лімфатичні вузли, що входять до складу передніх глибоких шийних лімфатичних вузлів.
Іннервують трубний мигдалик: чутлива інформація від мигдалика передається по мигдаликових гілках (rr. tonsillares) язико-глоткового нерва (ІХ черепний нерв), а також блукаючого нерва (Х черепний нерв); парасимпатичні завузлові волокна відходять від крило-піднебінного вузла (VII черепний нерв); симпатичні завузлові волокна відходять від зовнішнього сонного сплетення і проникають у трубний мигдалик разом з артеріальними гілками.
Лімфоїдні утворення в стінках травної, дихальної та сечової систем
Лімфоїдні вузлики (noduli lymphoidei). У власній пластинці слизової оболонки та в підслизовій основі стінок органів травної системи – глотки, стравоходу, шлунка, тонкої та товстої кишок, жовчного міхура; органів дихання – гортані, трахеї, великих бронхів; сечових органах – сечоводів, сечового міхура і сечівника – розташовані численні лімфоїдні вузлики (noduli lymphoidei). Їхня кількість складає близько 45 вузликів на 1 см2 площі слизової оболонки, які „слідкують” за антигенами, що проходять по цих шляхах. Крім того, в стінках цих органів міститься дифузна лімфоїдна тканина.
У стінках перелічених органів лімфоїдні вузлики містяться на різній відстані один від одного (від 1 мм до 5 мм) і на різній глибині. У слизовій оболонці лімфоїдні вузлики є численними структурами: у дітей в стінці тонкої кишки їх налічується понад 5000, а в стінці товстої кишки – приблизно 7000. У дорослої людини в стінках дванадцятипалої кишки і проксимального відділу порожньої кишки на 1 см2 поверхні слизової оболонки припадає 30–35 одиноких лімфоїдних вузликів, у дистальному відділі порожньої кишки
– 41–45 вузликів, а в клубовій кишці – 35–40 вузликів. Іноді лімфоїдні вузлики розміщені так близько до епітеліального покриву, що слизова оболонка над ними випинається у вигляді невеликих горбків. У слизовій оболонці гортані численні лімфоїдні вузлики розміщені у вигляді кільця, яке називають “гортанним мигдаликом”. Найбільше лімфоїдних вузликів міститься у слизовій оболонці задньої поверхні надгортанника і в бічних відділах присінка гортані, а також шлуночків гортані і черпакувато-надгортанних складок.
Розрізняють одинокі лімфоїдні вузлики (noduli lymphoidei solitarii) і скупчені лімфоїдні вузлики (noduli lymphoidei aggregati).
Одинокі лімфоїдні вузлики (noduli lymphoidei solitarii) мають кулясту або овоїдну форму діаметром 1,5–2 мм, їхня строма утворена ретикулярними клітинами і волокнами, серед яких розташовані Ті В-лімфоцити. У дитячому і юнацькому віці більшість лімфоїдних вузликів мають зародковий центр, або центр розмноження (centrum germinativum), в яких є багато лімфобластів, клітин, що мітотично поділяються, макрофагів і плазмоцитів (тобто відбувається активний лімфоцитопоез). Центр розмноження оточений темнішою крайовою, або плащовою зоною (zona marginalis) із щільно розташованих малих лімфоцитів. Навколо вузликів гемокапіляри утворюють сітку. Судини іннервують симпатичні завузлові волокна, що підходять до вузликів з відповідними артеріями. Частина активованих В-лімфоцитів мігрує у кровоносне русло через стінки закапілярних венул, інші – у тканини органа.
Скупчені лімфоїдні вузлики (noduli lymphoidei aggregati) складаються з 5 і більше (до 150) одиноких лімфоїдних вузликів, щільно розташованих один біля одного, і дифузної лімфоїдної тканини. У клубовій кишці наявна велика кількість скупчених лімфоїдних вузликів. Ці утворення називають також лімфоїдними бляшками, або бляшками Пейєра. Найчастіше вони овальної форми, довжина такої бляшки 1,5 см. Лімфоїдні бляшки містяться у власній пластинці слизової оболонки і в підслизовій основі стінки кишки. У цих місцях м’язова пластинка слизової оболонки відсутня. Слизова оболонка над бляшкою випинається і добре помітна. Розташовані бляшки переважно навпроти брижового краю кишки. У місцях, де розташовані лімфоїдні бляшки, слизова оболонка ворсинок не утворює, а колові складки перериваються. Довга вісь лімфоїдної бляшки переважно орієнтована вздовж кишки, хоча можливе поперечне або косе розміщення. Лімфоїдні бляшки розміщуються близько одна від одної, хоча інколи відстань між ними досягає кількох десятків сантиметрів. Слизова оболонка клубової кишки в ділянках лімфоїдних бляшок нерівна, горбиста, між горбками діаметром 1–2 мм є невеликі борозенки. Лімфоїдні вузлики в бляшках можуть розміщуватися двома рядами.
Одинокі лімфоїдні вузлики і скупчені лімфоїдні вузлики (лімфоїдні бляшки) починають утворюватись у стінках названих органів на 4–5-му місяці пренатального онтогенезу у вигляді скупчення клітин лімфоїдного ряду, а на 6–7-му місяці вже цілком сформовані. У немовлят бляшки клубової кишки ще не випинаються і мають довжину до 2 см, але у лімфоїдних вузликах вже наявні центри розмноження, таких бляшок є близько 30. У 12–15-річних підлітків кількість лімфоїдних бляшок у тонкій кишці зростає у декілька разів – до 122–316. Кількість великих бляшок довжиною понад 4 см у цьому віці досягає 9–12. Починаючи з юнацького віку число всіх лімфоїдних бляшок досягає 59–159, а в людей похилого і старечого віку – до 16–20. Кількість великих бляшок (довжиною понад 4 см) у них не перевищує шести. У 50–60-річних людей у лімфоїдних вузликах центри розмноження трапляються рідко, а в осіб похилого і старечого віку в бляшках розташована тільки дифузна лімфоїдна тканина.
Скупчені лімфоїдні вузлики червоподібного відростка (noduli lymphoidei aggregati appendicis vermiformis) розташовані у власній пластинці слизової облонки та підслизовій основі вздовж усього органа. Кількість лімфоїдних вузликів у стінці червоподібного відростка у дітей і підлітків є максимальною – до 550, а їх діаметр коливається в межах 0,2–1,2 мм, тому червоподібний відросток називають вторинним лімфоїдним органом, який контролює імунний статус кишкового тракту. Переважна більшість вузликів у цей період мають центри розмноження, в яких багато лімфобластів, клітин, що мітотично поділяються, макрофагів і плазмоцитів.
Лімфоїдні вузлики в стінці червоподібного відростка починають утворюватися у плода вже на 4-му місяці розвитку, спочатку у слизовій оболонці, а потім у підслизовій основі. Центри розмноження у лімфоїдних вузликах виникають перед народженням дитини або відразу після народження. Діаметр вузликів у цей період дорівнює 0,5–1,2 мм, а їхня кількість в органі вже становить 150–200. Починаючи з юнацького віку, кількість лімфоїдних вузликів зменшується; особливо інтенсивний цей процес у людей похилого і старечого віку. У червоподібному відростку значна частина лімфоїдних вузликів поступово замінюється жировою тканиною.