Курс лекцій з гістології
РОЗДІЛ 3 М’ЯЗОВІ ТКАНИНИ
М’язова тканина належить до високоспеціалізованих тканин, що забезпечують рух організму в просторі і скоротливі процеси у внутрішніх органах. Вона має досить різноманітну будову, характер скорочення і походить з різних ембріональних закладок. Все це створює необхідність розділяти м’язову тканину на різновиди. Можна класифікувати мускулатуру хребетних і безхребетних за різними ознаками:
- за будовою виділяють гладку, поперечносмугасту і мускулатуру з подвійною косою покресленістю;
- за розташуванням в організмі розрізняють мускулатуру внутрішніх органів, серцеву і соматичну;
- за функціями – тонічну, тетанічну і м’язову тканину з “замикальною” функцією;
- за походженням – ектодермальну, ентодермальну і мезодермальну.
Найбільше прийнятою класифікацією мускулатури хребетних є морфофункціональна, за якою розрізняють гладку і поперечносмугасту (скелетну і серцеву) м’язову тканину.
3.1. Гладка мускулатура
Гладка мускулатура хребетних тварин входить у м’язові оболонки судин, стінку шлунку, кишковика, сечового міхура, виявляється в селезінці, шкірі та інших органах. Структурною одиницею гладкої м’язової тканини служить гладком’язова клітина – міоцит. Вона має, як правило, веретеноподібну форму. Довжина клітин коливається від 20 до 500 мкм, а поперечник рівний 10–20 мкм. Найбільша величина гладком’язових клітин спостерігається в матці під час вагітності.
У розширеній центральній частині клітини розташовується подовжене ядро зі щільно упакованим пристінково розподіленим хроматином. З поверхні м’язова клітина оточена мембраною – міолемою, тотожною плазмолемі інших клітин. Ззовні вона вкрита ще тонкою базальною мембраною (пластинкою), до зовнішньої поверхні якої прикріплюються колагенові і аргирофільні волокна. Окрім механічної функції, базальні пластинки, які оточують клітини, виконують важливу роль в підтримці клітинного гомеостазу.
Клітини можуть утворювати у внутрішніх органах пласти або тяжі великої протяжності, об’єднані сполучнотканинними прошарками і пронизані судинами і нервами. Нервові закінчення, звичайно, підходять не до кожної м’язової клітини, а до групи з 10–12 клітин, що є ефекторною одиницею.
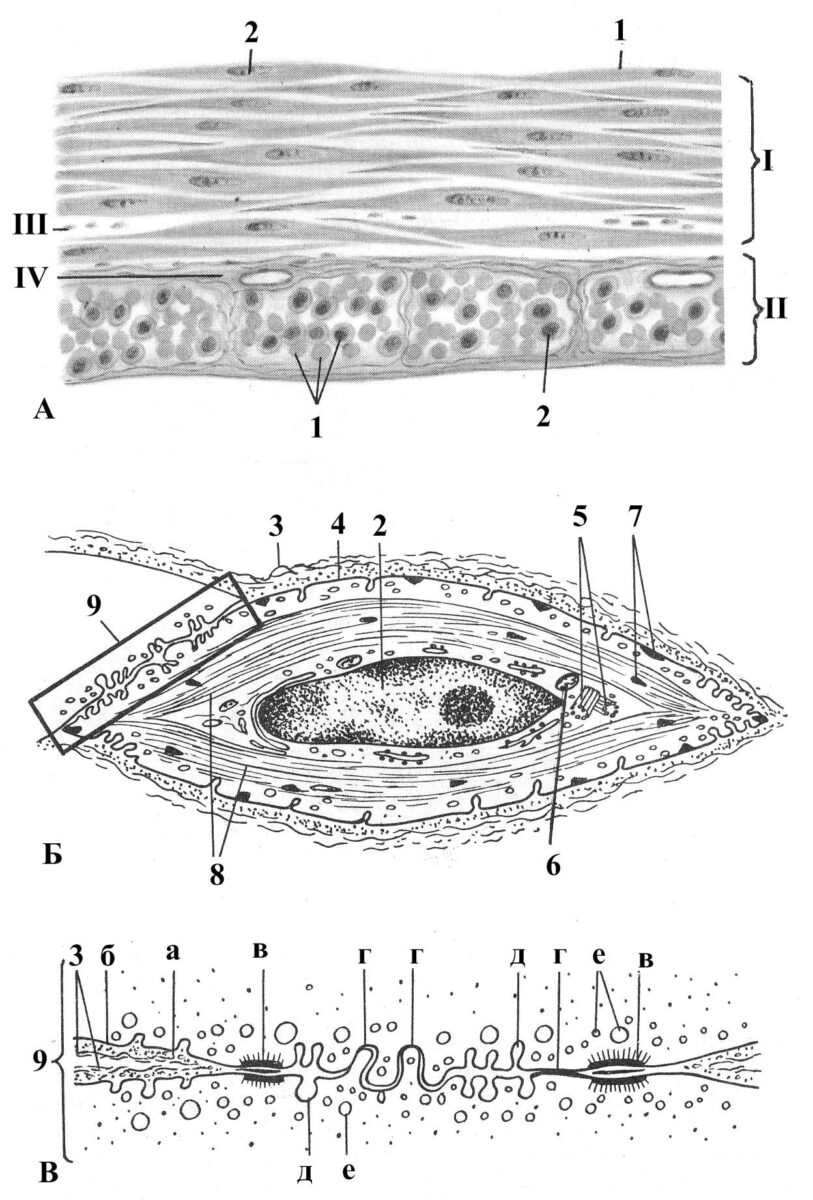
Рис. 3.1. Гладка м’язова тканина. А – препарат, Б – ультраструктура гладкого міоциту, В – зона міо-міоцитного контакту (схеми). І – повздовжній та ІІ – поперечний шари гладких міоцитів; ІІІ – ендомізій; ІV – перимізій. 1 – гладкий міоцит; 2 – ядро; 3 – еластичні волокна; 4 – міолемма; 5 – клітинний центр; 6 – мітохондрії; 7 – щільні тільця; 8 – міофіламенти; 9 0 міо-міоцитний контакт: а – базальна мембрана, б – плазмолемма, в – десмосома, г – нексус, д – кавеоли, е – везикули.
Електронно-мікроскопічні дослідження гладком’язової клітини показали, що міофібрил, тобто мікроскопічно видимих ниток, що тягнуться через всю клітину, і описаних на рівні світлової мікроскопії, насправді не існує.
У цитоплазмі цих клітин присутні лише подовжньо орієнтовані субмікроскопічні міофіламенти, звичайно зібрані в пучки. Довжина міофіламентів не перевищує 1–2 мкм. У тонких міофіламентах міститься білок актин, а в товстих – міозин (у скороченому м’язі). У цитоплазмі гладком’язових клітин знаходяться у великій кількості дрібні міхурці, які містять іони кальцію, необхідні для здійснення скорочення. Ці міхурці за своїм призначенням, очевидно тотожні елементам саркоплазматичного ретикулуму поперечносмугастих м’язів. Разом зі структурами, безпосередньо пов’язаними зі здійсненням скорочення, у гладком’язовій клітині є “трофічні” компоненти, до них належать мітохондрії, внутрішньоклітинний сітчастий апарат, клітинний центр, ендоплазматичний ретикулум, включення глікогену. Всі вони розташовуються, в основному, поблизу ядра і забезпечують обмін речовин у клітині.
Гладка мускулатура має здібність до фізіологічної репаративної і внутрішньоклітинної регенерації.
3.2. Скелетна мускулатура
Скелетна м’язова тканина будується з поперечносмугастих м’язових волокон. Структурною одиницею цієї тканини служить не клітина, як у гладкій мускулатурі, а поперечносмугасте волокно – симпластичне багатоядерне утворення. Волокна мають витягнуту циліндричну форму. Їх довжина варіює від 400–600 мкм до 10–12 см, а поперечник від декількох десятків до сотень мікрон. Кожне м’язове волокно покрите оболонкою – сарколемою. Вона складається з двох основних шарів: внутрішнього – плазмолеми, подібній плазматичній мембрані інших клітин, і зовнішнього – базальної мембрани, що містить в собі тонкі волокнини, склеєні аморфною речовиною, і тісно спаюється з колагеновими і аргирофільними волокнами навколишньої сполучної тканини.
Кожне м’язове волокно, таким чином, оточене тонким прошарком сполучної тканини, яку називають ендомізієм. Групи м’язових волокон утворюють пучки, покриті міцнішою сполучнотканинною оболонкою – перимізієм. Увесь м’яз, в цілому, охоплюється фасцією, або епімізієм. По сполучнотканинних прошарках усередині м’яза проходять кровоносні судини, що розгалуджуються на капіляри, а також нерви, які утворюють на м’язових волокнах рецепторні й афекторні закінчення. Розподіл і кількість ядер може бути різним. Навколо ядер і між міофібрилами знаходиться саркоплазма (цитоплазматичні м’язові волокна) з різними органоїдами і включеннями.
Скоротливими елементами поперечносмугастого м’язового волокна є міофібрили, які заповнюють основну його масу. Поперечник міофібрил рівний 0,5–2 мкм, а довжина зрівнюється з довжиною всього волокна.
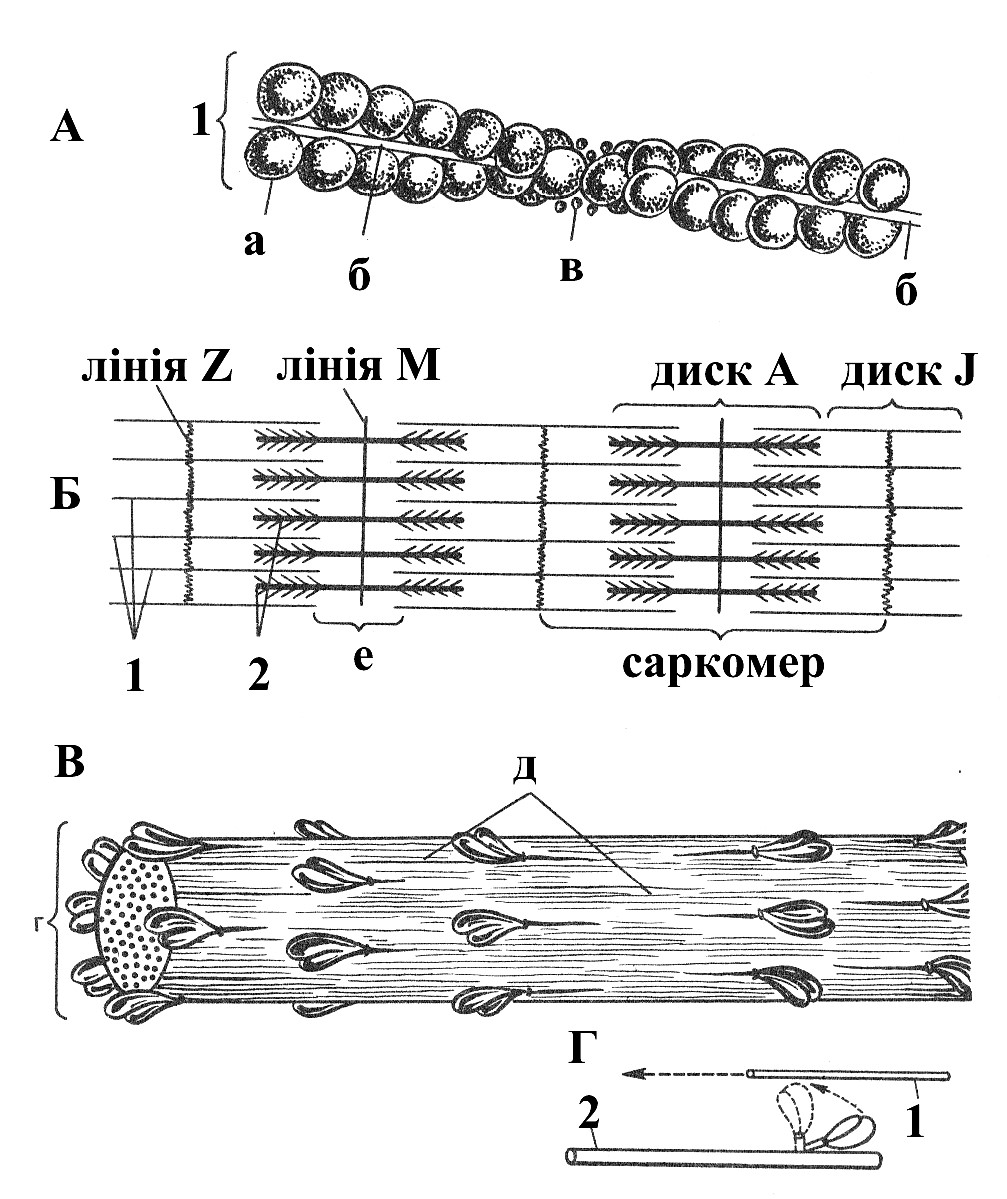
Рис. 3.2. Будова міофібрили. А – склад та структура тонкого міофіламента. Б – порядок розташування міофіламентів у міофібрілі. В – склад та структура товстого міофіламенту. Г – принцип «роботи» поперечного мостика (схеми). 1 – тонкий міофіламент: а – мономер актину, б – тропоміозин, в – тропонін; 2 – товстий міофіламент: г – шість рядів поперечних мостиків – голівки молекул міозину, д – хвостові ділянки молекул міозину, е – світла зона.
Поперечна покресленість міофібрил пов’язана з їх фізичною і хімічною неоднорідністю. У міофібрилах розрізняють темні анізотропні ділянки з подвійним променезаломленням (А-диски) та ізотропні світлі ділянки без подвійного променезаломлення (I-диски), закономірно розташовані уподовж волокна. У розслабленому м’язі в центрі кожного ізотропного диска знаходиться «смужка Z». У середній частині анізотропного диска помітна світла зона – диск Н, в самому центрі якого розташована «смужка М» (мезофрагма). За рахунок чергування ізотропних й анізотропних сегментів кожна міофібрила набуває поперечної покресленості. Оскільки у волокні міофібрили розташовуються впорядковано, так, що ізотропні й анізотропні диски сусідніх міофібрил точно співпадають – все волокно, в цілому, також має поперечну покресленість.
Структурною одиницею міофібрили служить саркомер – ділянка міофібрили, що обмежена двома смужками Z і складається з половини ізотропного, цілого анізотропного і половини наступного ізотропного диска.
Кожна міофібрілла складається з паралельно лежачих ниток або протофібрил різної товщини (“товстих і тонких”). Поперечник тонких – 50– 70 Å, товстих – 100–120 Å. На поперечних зрізах м’язового волокна протофібрили мають вид дрібних і крупних крапок, упакованих гексагонально. Кожна товста протофібрила оточена шістьма тонкими, причому співвідношення товстих ниток до тонких складає 1:2.
Тонкі нитки, створюючі ізотропні диски, містять білок актин. Одним своїм кінцем ці нитки прикріплюються до Z-смужки, що перетинає І-диск посередині. Товсті нитки містять міозин і представляють основну частину Адисків. У нескороченому саркомері вільні кінці актинових ниток заходять між міозиновими, закінчуючись біля меж дисків Н. Таким чином, у периферичних ділянках А-дисків присутні й міозинові, й актинові нитки, які в цих ділянках перекривають одна одну (“зона перекриття”). У цих зонах між товстими і тонкими протофібрилами виявляються короткі поперечно орієнтовані містки довжиною 130 Å, які розташовані на відстані 143 Å по вісі волокнини.
У поперечносмугастому м’язовому волокні можна розрізнити скоротні, трофічні й опорні структури. До перших належать міофібрили, до других – компоненти цитоплазми, які забезпечують метаболізм, до третіх – смужки Z, М, канали Т-системи, що упорядковують розташування міо- і протофібрил, а також базальна мембрану і плазмолема з її різними диференціюваннями, що здійснюють міцний зв’язок м’язового волокна із сухожиллям.
Скорочення міофібрил. У процесі скорочення міофібріли відбувається взаємне ковзання тонких ниток відносно товстих, ніби втягування ізотропного диска в анізотропний, внаслідок чого довжина саркомеру зменшується. Число поперечних містків між товстими і тонкими нитками при цьому збільшується. У скороченому саркомері міозинові протофібрили рухаються до зіткнення зі смужками Z, а актинові нитки, рухаючись назустріч одна одній, досягають середини анізотропних дисків або навіть заходять за рівень смужки М, тобто перекривають одна одну.
Вважають, що в області актинового центру міозину є короткі ділянки поліпептидного ланцюжка, які можуть змінювати свою конфігурацію, то витягуватись, то скручуватись в α-спіраль з утворенням водневих зв’язків. Коли міофібрила знаходиться у розслабленому стані, в середовищі іони Са2+ відсутні і зберігається, хоча і безладна, але витягнута конфігурація поліпептидного ланцюжка-“містка”.
При активації міофібріли іони Са2+, що поступають із саркоплазматичного ретикулуму, сполучають негативний іон АДФ, прикріплений до актину, з іоном АТФ кінця містка. Негативний заряд АТФ у результаті цього нейтралізується, сила відштовхування зникає і поліпептидний ланцюжок коротшає, частково перебудовуючись в α-спіраль.
Такий процес супроводжується звільненням потенційної енергії, яка була запасена у витягнутому поліпептидному ланцюжку. При вкороченні цього ланцюжка відбувається переміщення актинової нитки відносно міозинової на один “крок” у напрямку до центру саркомера. Останній при цьому коротшає.
В результаті вкорочення містка АТФ, приєднаний до кінця поліпептидної нитки, наближається до області АТФ-азного центру цього ланцюга і відбувається відщеплювання останньої фосфатної групи від АТФ”містка”, тобто АТФ перетворюється в АДФ. Зв’язок між актиновою і міозиновою протофібрилами при розщепленні АТФ порушується. Можна сказати, що даний міозиновий місток відпрацював.
Для підготовки до нового циклу роботи містка необхідна “підсадка” нового іона АТФ замість того, що утворив АДФ, що і відбувається, мабуть, за рахунок заміни цього АДФ на АТФ цитоплазми. Поява АТФ на кінці містка призводить до нового його розтягування. Місток при цьому готовий до повторного циклу роботи. Така робота відбувається, якщо в середовищі наявні іони Са2+. Після припинення збудження, тобто в процесі розслаблення м’яза, іони Са2+ віддаляються в саркоплазматичний ретикулум, так що всі містки виявляються витягнутими, але не входять у контакт з актиновими нитками.
Для роботи м’язового волокна дуже велике значення має саркоплазматичний ретикулум. У його цистернах (особливо літеральних, прикордонних з поперечними каналами ділянках) відбувається накопичення Са2+, таких важливих для акту скорочення. Крім того, саркоплазматичний ретикулум є джерелом швидкого отримання енергії, необхідної для скорочення міофібрил.