Курс лекцій з гістології
Гранулоцити (зернисті лейкоцити)
Нейтрофільні гранулоцити, нейтрофіли мають округлу форму, їх діаметр в краплині крові близько 7–9 мкм. У крові дорослої людини нейтрофілів міститься більше, ніж інших лейкоцитів; їх кількість досягає 65– 75 % від загального числа лейкоцитів. Залежно від ступеня диференціювання і форми ядра розрізняють сегментоядерні нейтрофіли – зрілі клітини з ядром, що складається з 3–5 сегментів, сполучених тонкими перемичками, і паличкоядерні нейтрофіли – незрілі форми клітин з підковоподібним ядром. Цитоплазма нейтрофілів слабооксифільна, в ній міститься дрібна зернистість. Гранули нейтрофілів є спеціалізованими лізосомами, що містять не менше 10 білків-ферментів, які знищують мікроорганізми: катепсини, еластазу, протеїнази – для розщеплення білків, а також лізоцим – для руйнування клітинної оболонки бактерій. Нейтрофіли – рухомі клітки, вони можуть мігрувати з кровоносних судин і пересуватися до джерела подразнення. Нейтрофіли мають високу здатність до фагоцитозу і беруть участь у гострих запальних реакціях. Головна їх функція – руйнування і фагоцитоз тканинних уламків і мікроорганізмів. Тривалість життя нейтрофілів близько 8 діб, при цьому в кров’яному руслі вони знаходяться 8–12 годин, а потім виходять у сполучну тканину, де виявляється їх максимальна активність. Нейтрофіли містять мітохондрії і велику кількість глікогену. Клітина одержує енергію шляхом гліколізу, що дозволяє їй існувати у бідних киснем пошкоджених тканинах. Кількість органел, необхідних для синтезу білка, мінімальна, тому нейтрофіл не здатний до тривалого функціонування і гине після єдиного спалаху активності. Загиблі нейтрофіли складають основний компонент гною (гнійні клітини). До складу гною також входять загиблі клітини сполучної тканини, бактерії, тканинна рідина.
Базофільні гранулоцити, базофіли мають діаметр близько 9 мкм у краплині крові. У крові людини вони складають 0,5–1% від загального числа лейкоцитів. Знаходяться в межах кісткового мозку і кровоносного русла, в крові циркулюють 1–2 діб. Як і інші лейкоцити, вони можуть покидати кровотік, але їх здатність до амебоїдного руху обмежена. Частіше мають тридольне S-подібне ядро. Містять всі види органелл, вільні рибосоми, глікоген.
Цитоплазма базофілів заповнена крупними овальними гранулами з щільним вмістом.
Функція базофілів полягає в їх участі в метаболізмі гістаміну й гепарину, які необхідні для регуляції процесів зсідання крові та проникності судин. Базофіли беруть участь в імунологічних реакціях організму, зокрема в реакціях алергічного характеру. Базофіли реагують на комплекс антиген– антитіло. Вони легко приєднують особливі антитіла – імуноглобуліни Е (IgE), що циркулюють у крові. Чужорідні білки-антигени (алергени) спричинюють утворення IgE-антитіл, які їх зв’язують. Комплекси IgE+антиген, діючи на базофіли, призводять до їх дегрануляції і вивільненню гістаміну. Це обумовлює різке розширення судин, появу набряків тощо.
Фагоцитарна активність базофілів виражена слабо.
Еозинофільні гранулоцити, еозинофіли – крупніші клітини, в порівнянні з нейтрофільними, діаметр 9–10 мкм. Їх кількість коливається в межах 1–5% від загального числа лейкоцитів. Термін знаходження еозинофілів у кров’яному руслі – 3–8 годин, у подальшому вони мігрують у тканини, що контактують із зовнішнім середовищем (слизові оболонки дихальних і сечостатевих шляхів, кишковика), де продовжують функціонувати.
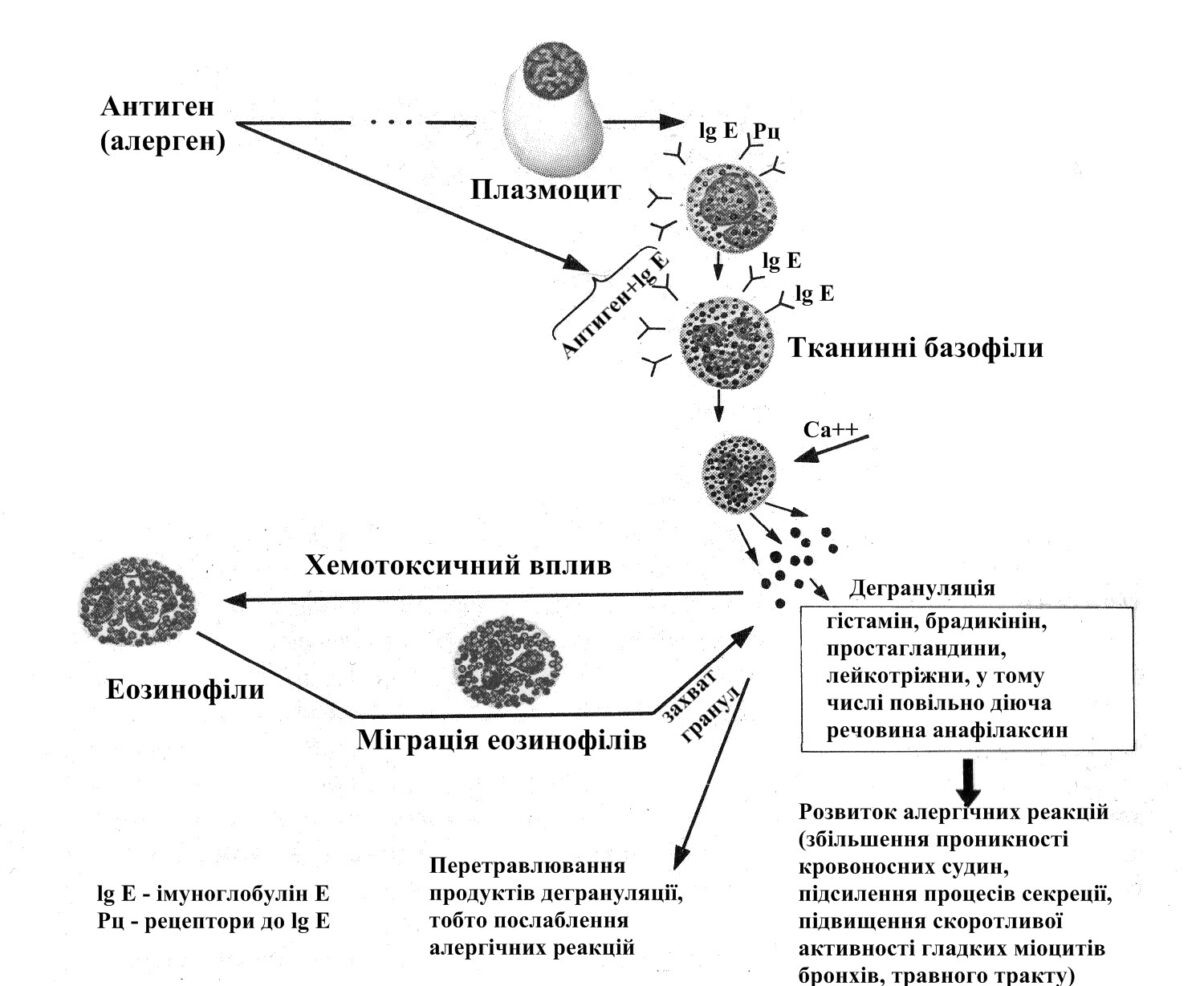
Рис. 2.3. Схема участі еозинофільних та базофільних гранулоцитів в алергічних реакціях
Залежно від стадії розвитку розрізняють сегментоядерні еозинофіли – зрілі клітини з ядром, що складається з двох крупних сегментів, сполучених тонкою перемичкою, і паличкоядерні еозинофіли – незрілі клітини з підковоподібним ядром. Цитоплазма містить добре розвинену гранулярну ендоплазматичну мережу, невелику кількість цистерн гладкої ендоплазматичної мережі, скупчення рибосом, окремі мітохондрії, багато глікогену. Характерна ознака еозинофілів – наявність специфічних оксифільних гранул овальної форми, в яких містяться протеолітичні ферменти, а також гістаміназа, фосфоліпаза D.
Еозинофіли здатні до фагоцитозу, проте їх фагоцитарна активність нижча, ніж у нейтрофілів. Вони беруть участь у захисних реакціях організму на чужорідний білок у алергічних і анафілактичних реакціях. Виявлено, що в областях розвитку реакції антиген–антитіло у вогнищах запалення утворюються речовини, які стимулюють вихід еозинофілів з кісткового мозку в циркулюючу кров і звідти в тканини. Еозинофіли мають позитивний хемотаксис до пептиду, що виділяється тканинними базофілами при руйнуванні їх гранул. Ферменти еозинофілів руйнують речовини, що вивільняються базофілами при імунних реакціях (алергія, анафілаксія): арілсульфатаза руйнує анафілаксин, гистаміназа – гістамін.
Не маючи здатності синтезувати гістамін, еозинофіли можуть накопичувати його, фагоцитуючи гістаміновмісні гранули, які виділяються базофілами, а також абсорбувати його на своїй цитолемі за допомогою специфічних рецепторів до гістаміну. Крім того, еозинофіли виробляють спеціальний чинник – сигнальну речовину, що гальмує вивільняння гістаміну з базофілів.
Функцією еозинофілів також є знищення паразитів у місцях їх занурення в тканини організму (але не в області їх остаточної локалізації). Специфічні цитоплазматичні гранули еозинофілів містять набір протеолітичних і гідролітічних ферментів та подовжений кристалоїд, що має кубічну структуру і складається з антипаразитарного агента (багатого аргініном лужного білка), лізосомних ферментів і пероксидази. Після активації антитілами і компонентами комплементу еозинофіли виділяють вміст гранул і ліпідні медіатори, що ушкоджують оболонки паразитів. Секреція вмісту гранул запускається протягом декількох хвилин і може продовжуватися декілька годин. Після дегрануляції еозинофіли знищуються апоптозом.
Термін життя еозинофілів приблизно 8–14 днів. Вміст їх у крові коливається протягом доби (максимум – у нічні години, мінімум – у ранкові). Агранулоцити (незернисті лейкоцити)
Лімфоцити у крові дорослих людей складають 20–35%. Залежно від розмірів та стадії диференціювання розрізняють великі, середні та малі (найбільше зрілі) лімфоцити.
Малі лімфоцити поділяються в свою чергу на світлі і темні, складають велику частину (близько 70–75%) всіх лімфоцитів крові людини.
Середні – 10–12%, мають діаметр близько 10 мкм.
Плазмоцити (лімфоплазмоцити) складають у крові людини 1–2%.
Серед лімфоцитів по шляхах диференціювання і ролі в становленні захисних реакціях виділені два основні види – Т- і В-лімфоцити.
Т-лімфоцити (тимусзалежні лімфоцити) утворюються зі стовбурових клітин кісткового мозку в тімусі, вони забезпечують реакції клітинного імунітету і регуляцію гуморального імунітету.
Це лімфоцити-довгожителі, можуть жити кілька (десятки) років. Серед популяції Т-лімфоцитів розрізняють кілька субпопуляцій: Т-кілери, або клітини-вбивці, специфічний цитотоксичний ефект яких забезпечує протипухлинний і трансплантаційний імунітети; Т-хелпери (помічники) мають здатність специфічно розпізнавати антиген і посилювати утворення антитіл В-лімфоцитами; Т-супресори пригнічують здатність В-лімфоцитів до продукції антитіл; Т-клітини пам’яті – лімфоцити, що довгий час зберігають інформацію про антиген.
В-лімфоцити, або бурсазалежні, утворюються у птахів у фабрицієвій сумці. А у людини В-лімфоцити утворюються зі стовбурових клітин у кістковому мозку, подальше їх диференціювання відбувається у лімфатичних фолікулах шлунково-кишкового тракту. Їх головна функція – синтез захисних білків-імуноглобулінів (антитіл) і забезпечення гуморального імунітету. В-лімфоцити здатні перетворюватись у плазмоцити, які також продукують антитіла.
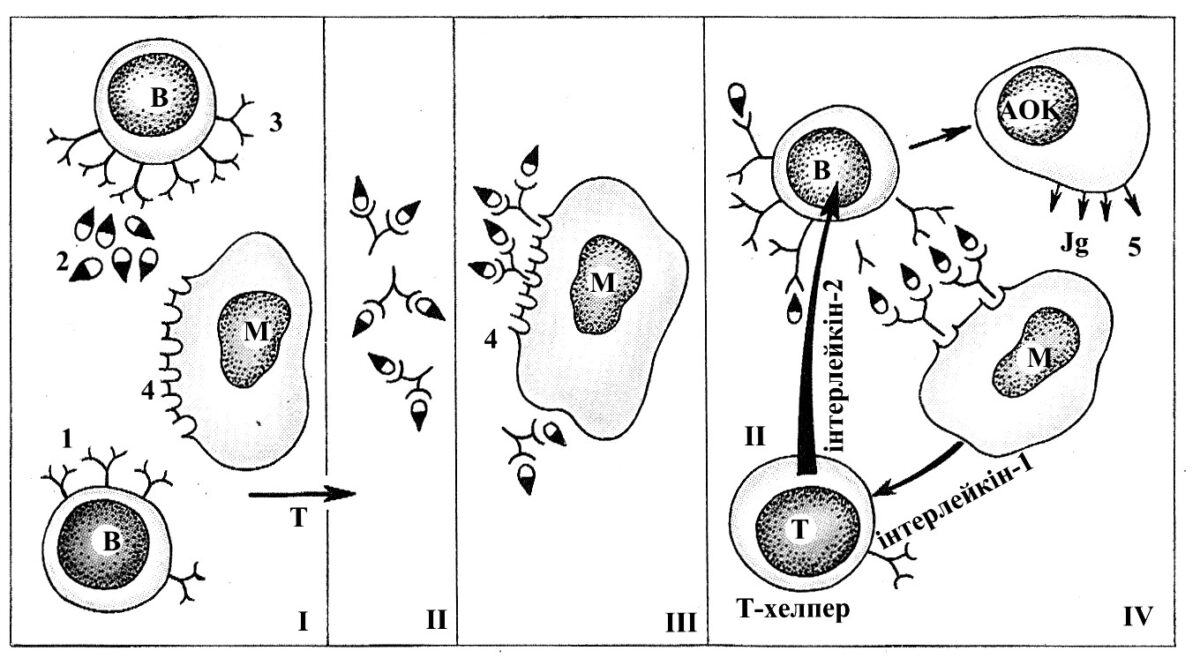
Рис. 2.4. Взаємодія клітин в імунній реакції. Схема взаємодії Т- і Влімфоцитів та макрофагів. І – антиген-реактивні Т-лімфоцити за допомогою своїх рецепторів розпізнають чужорідні молекули антигенів; ІІ-ІІІ – комплекси рецепторів Т-лімфоцитів та антигенів захоплюються та перетравлюються макрофагами, які мають власні рецептори; ІV – Влімфоцити, контактуючи з макрофагами, за допомогою свої рецепторів отримують від них антигенові інформацію. Т-лімфоцит-хелпер продукує медіатор – «індуктор імцнопоезу» (інтерлейкін-2). В результаті цих взаємодій В-лімфоцит трансформується в клітини, які продукують антитіла, інактивуючі антигени.
Т – Т-лімфоцити; М – макрофаги; ІІ – «індуктор імунопоезу»; АОК – Клітини, які продукують антитіла. 1 – рецептори Т-лімфоцитів; 2 – антиген; 3 – рецептори В-лімфоцитів; 4 – рецептори макрофагів; 5 – антитіла.
Виявляють Т- і В-лімфоцити та їх субпопуляції імунологічними методами, що базуються на специфічності будови мембран цих клітин.
Моноцити набагато крупніші за інші лімфоцити (9-12 мкм) в краплині крові, їх кількість у крові людини коливається в межах 6–8% від загального числа лейкоцитів. За діаметром це найбільші клітини серед лейкоцитів. У цитоплазмі містяться всі органели, численні лізосоми. Ядро найчастіше бобовидне. Моноцити належать до макрофагічної системи організму.
Клітини цієї системи характеризуються походженням з промоноцитів кісткового мозку, здатністю до піноцитозу й імунного фагоцитозу.
Час перебування моноцитів у крові варіює від 36 до 104 годин, після чого вони виходять із судин і в тканинах перетворюються на тканинні макрофаги, які є кінцевою стадією диференціювання цих клітин крові.
Тромбоцити (кров’яні пластинки) мають вид найдрібніших безбарвних тілець округлої, овальної, веретеноподібної або неправильної форми, що відокремилися від гігантських клітин кісткового мозку – мегакаріоцитів. Тромбоцити – без’ядерні фрагменти їх цитоплазми розміром 2–3 мкм.
Завдяки здатності до склеювання на препаратах крові вони звичайно зустрічаються групами.
Кров’яні пластинки беруть участь у процесі зсідання крові. Ця функція визначається їх здатністю швидко розпадатися, склеюватися в групи, навколо яких виникають нитки фібрину. В процесі зсідання крові кров’яні пластинки виділяють низку речовин (фосфоліпіди, ліпопротеїди та ін.) та різні ферменти (каталаза, кисла фосфотаза тощо).
Тривалість життя кров’яних пластинок близько 5–8 днів.
Кров’яні пластинки ссавців у неактивному стані мають форму круглих дисків, підтримуваних периферичним кільцем мікротрубочок. Поверхня пластинок утворює систему вп’ячувань (відкритих канальців), які можуть далеко заглиблюватися у цитоплазму.
Мембрана пластинок оснащена різноманітними рецепторами: до тромбіну, АДФ (регулятору агрегації пластинок), катехоламінам, колагену і т.д. Дуже добре розвинений субмембранний скоротливий апарат, або система мікрофіламентів. Мікрофіламенти – це комплекс актина з нем’язовим міозином. При активації пластинок мережа філаментів скорочується, стягуючи органели до центру: відбувається централізація органел.
Оскільки тромбоцити – елементи крові з активними метаболічними процесами, в їх цитоплазмі багато мітохондрій. Енергетику пластинок також забезпечують відкладення глікогену. Характерними органелами тромбоцитів є канальця, заповнені електронно-щільним вмістом (так звана щільна тубулярна система). Ці канальця – різновид ендоплазматичної мережі, вони накопичують іони Са++, які виходять в цитоплазму у момент скорочення пластинок, а скорочення – це Са-залежний процес. Канальці тубулярної системи активно контактують з поверхневими відкритими канальцями.
Іншою функцією щільних канальців, ймовірно, є синтез простагландинів – регуляторів скорочення судин.
Найбільше специфічні органели тромбоцитів – гранули, серед яких розрізняють α-гранули (помірної густини) і β-гранули, або щільні тільця. У число α-гранул, ймовірно, входять звичні лізосоми і пероксисоми. У складі α-гранул знайдений цілий спектр речовин, серед яких – гідролітичні ферменти, чинники зсідання крові (специфічні чинники тромбоцитів, АДФ, АТФ) і скорочення судин (серотонін), фібриноген, тромбоцитарний чинник росту, Са2+ і т.д. У β-гранулах містяться, головним чином серотонін, катехоламіни, АДФ і АТФ. Серотонін і катехоламіни поглинаються пластинками з плазми крові.
Хоча тромбоцити – фрагменти клітин, вони проявляють активність, властиву справжнім клітинам: здатність до розпластування, скорочення, пересування (зокрема, хемотаксису), піноцитозу. Фагоцитуватиь вони не можуть, хоча чужорідні частинки адсорбуються на їх поверхні і проникають вглиб відкритих канальців.
Сигналом до активації пластинок є взаємодія з тромбіном, АДФ або колагеном.
Гемограма. Лейкоцитарна формула. При клінічних аналізах досліджують хімічний склад крові, визначають кількість еритроцитів, лейкоцитів, гемоглобіну, швидкість осідання еритроцитів (ШОЕ). У здорової людини формені елементи крові знаходяться у певних кількісних співвідношеннях, які прийнято називати гемограмою, або формулою крові. Важливе значення для характеристики стану організму має диференційне підрахування лейкоцитів. Відсоткові співвідношення різних форм лейкоцитів називають лейкоцитарною формулою.
Лімфа це жовтувата рідина білкової природи. Складається вона з лімфоплазми і формених елементів. За хімічним складом лімфоплазма близька до плазми крові, але містить менше білка, альбуміни переважають над глобулінами.
Формені елементи лімфи представлені лімфоцитами (98%), а також моноцитами та іншими видами лейкоцитів. Лімфа утворюється в лімфатичних капілярах тканин і органів, куди під впливом різних чинників, зокрема осмотичного і гідростатичного тиску, з тканин постійно надходять різні компоненти лімфоплазми. З капілярів лімфа переміщається в периферичні лімфатичні судини, по них – в лімфатичні вузли, потім у крупні лімфатичні судини і вливається у кров. Склад лімфи постійно змінюється. Розрізняють периферичну лімфу (до лімфатичних вузлів), проміжну (після проходження через лімфатичні вузли) і центральну (лімфу грудної і правої лімфатичних проток). Процес лімфоутворення тісно пов’язаний з надходженням води та інших речовин з крові в міжклітинні простори і утворенням тканинної рідини.