Периферійна частина нервової системи
Нервову систему людини поділяють на центральну і периферійну частини. Перша включає спинний і головний мозок, друга складається із периферійних нервів .
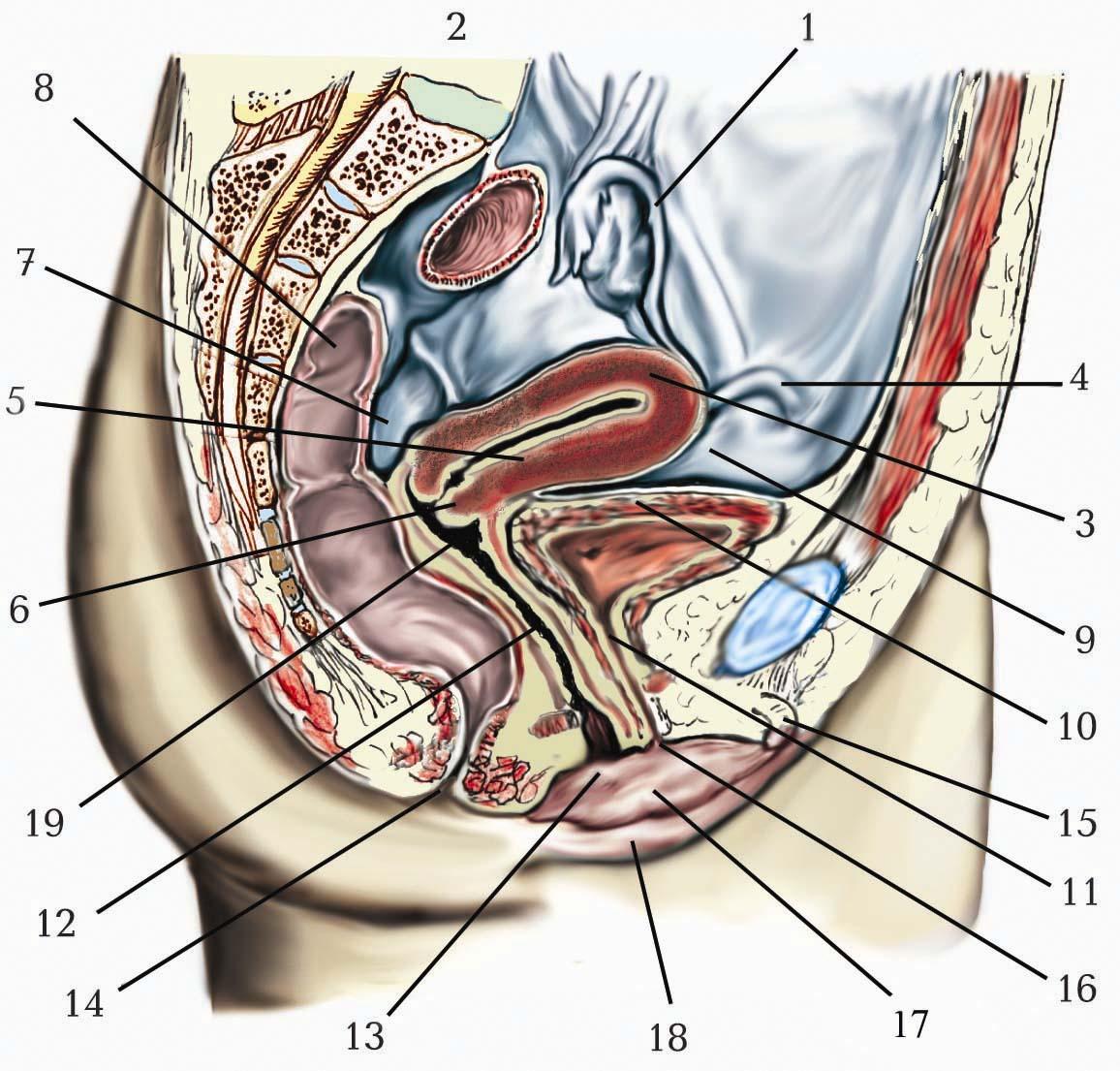
Периферійна частина нервової системи (периферійна нервова система) представляє собою сукупність спинномозкових і черепних нервів. До неї належать вузли та сплетення, що утворюють нерви, а також чутливі та рухові закінчення нервів. Окремо виділяють автономний відділ (автономну частину) периферійної нервової системи, що складається з симпатичної та парасимпатичної частин . Таким чином, периферійна нервова система об’єднує всі нервові утворення поза спинним і головним мозком. Таке об’єднання умовне, оскільки еферентні волокна, що входять до складу периферійних нервів, є відростками нейронів, тіла яких містяться в ядрах спинного і головного мозку. З функціональної точки зору периферійна частина нервової системи складається з провідників, що сполучають нервові центри з рецепторами і робочими органами. Анатомія периферійних нервів має велике значення для клініки, як основа для діагностики та лікування захворювань та уражень цього відділу нервової системи.
ПЕРИФЕРІЙНА ЧАСТИНА НЕРВОВОЇ СИСТЕМИ
Периферійні нерви
Чeрепні нерви (nervi craniales) належать, як і спинномозкові нерви, до периферійної частини нервової системи. Вони іннервують шкіру, м’язи, залози, внутрішні органи в ділянці голови. Деякі з них мають значну протяжність і іннервують внутрішні органи, які розташовані в ділянці шиї, грудей і живота. Крім того, черепні нерви є нервами органів чуття.
Черепні нерви впливають на стан різних відділів головного мозку, оскільки беруть участь у здійсненні мовлення, міміки, поведінки тощо. Багато з них пов’язані з зоровим, слуховим, смаковим, нюховим, шкірним, пропріоцептивним та інтероцептивним аналізаторами, є частиною складних кірково-ретикулярно-стовбурових чи рецепторно-стовбурово-таламокіркових функціональних систем. Існуючі анатомофункціональні зв’язки між черепними нервами і лімбічно-ретикулярним комплексом відіграють величезну роль у регуляції внутрішнього середовища організму, у тому числі залежно від оточуючого середовища.
Розрізняють 12 пар черепних нервів, кожна з яких має власний номер і назву:
І пара – нюховий нерв (nervus olfactorius);
II пара – зоровий нерв (nervus opticus);
ІІІ пара – окоруховий нерв (nervus oculomotorius);
IV пара – блоковий нерв (nervus trochlearis); V пара – трійчастий нерв (nervus trigeminus); VI пара – відвідний нерв (nervus abducens); VII пара – лицевий нерв (nervus facialis);
VIII пара – присінково-завитковий нерв (nervus vestibulocochlearis);
ІХ пара – язико-глотковий нерв (nervus glossopharyngeus);
Х пара – блукаючий нерв (nervus vagus);
ХІ пара – додатковий нерв (nervus accessorius); ХІІ пара – під’язиковий нерв (nervus hypoglossus). Номер черепного нерва свідчить:
1) про ростро-каудальне упорядкування, згідно з яким кожний нерв виходить через отвори в основі черепа;
2) про ростро-каудальне упорядкування, згідно з яким справжні черепні нерви прикріплюються до стовбура головного мозку (винятком є ХІ пара, що прикріплюється до рострального кінця спинного мозку). Назва нерва дає відомості щодо його будови, топографії або функції, але не має великого класифікуючого значення.
Черепні нерви поділяють на:
1) несправжні (І–ІІ пари);
2) справжні (ІІІ–ХІІ пари) нерви.
Несправжні черепні нерви (І–ІІ пари) є виростками переднього мозку: І пара – нюхового мозку; ІІ пара – проміжного мозку. Тому мієлін цих нервів має походження з олігодендроглії, на відміну від мієліну справжніх нервів, який утворюють клітини Шванна. Демієлінізуючі захворювання ЦНС (такі як розсіяний склероз) призводять до ураження тільки несправжніх нервів. І навпаки, захворювання справжніх нервів не поширюються на нюховий і зоровий нерви. Таким чином, терміни нюховий і зоровий “нерви” – є неправильними з точки зору ембріології, гістології та патології.
Справжні черепні нерви поділяють на:
1) чутливі нерви (VIII пара), які містять тільки чутливі волокна і відповідають (гомологічні) задньому корінцю спинномозкового нерва;
2) рухові нерви (III, IV, VI, XI, XII пари), які містять тільки рухові або рухові та вегетативні парасимпатичні волокна і відповідають (гомологічні) передньому корінцю спинномозкового нерва;
3) змішані нерви (V, VII, IX, X пари), у складі яких проходять як чутливі, так і рухові волокна і які подібні до спинномозкових нервів.
Коли справжній черепний нерв містить чутливі волокна, тоді він обов’язково має чутливий вузол черепного нерва (ganglion sensorium nervi сranіalis) із чутливими псевдоуніполярними або біполярними нейронами, що знаходиться поза мозком і відповідає чутливому вузлу спинномозкового нерва (ganglion sensorium nervi spinalis); а також чутливі центри – ядра (nuclei) у стовбурі головного мозку, що відповідають чутливим ядрам задніх рогів спинного мозку.
Коли справжній черепний нерв містить рухові волокна, він має рухові ядра у стовбурі головного мозку, що відповідають руховим ядрам передніх рогів спинного мозку.
Коли у складі справжнього черепного нерва проходять парасимпатичні нервові волокна, він має парасимпатичні ядра у стовбурі головного мозку. За ходом такого черепного нерва обов’язково повинен бути парасимпатичний вузол (ganglion parasympathicum) з мультиполярними нейронами, в якому відбувається переключення парасимпатичних волокон.
Несправжні черепні нерви є чутливими, але не мають чутливих ядер і чутливих вузлів.
Особливості філота онтогенезу черепних нервів пов’язані з такими факторами:
1) розвиток органів чуття і зябрових дуг;
2) редукція головних сомітів.
III, IV, VI пари черепних нервів пов’язані з середнім мозком і мостом, а також з головними (передвушними) міотомами. ІІІ пара черепних нервів пов’язана з І передвушним міотомом; IV пара черепних нервів – з ІІ міотомом; VI пара черепних нервів – з ІІІ міотомом.
V,VII, VIII, ІХ, Х, ХІ пари черепних нервів пов’язані з ромбоподібним мозком і зябровими дугами. Це нерви зябрового походження. V пара черепних нервів пов’язана з І зябровою дугою; VII i VIII (яка відділилася від VII в процесі розвитку) пари черепних нервів – з ІІ зябровою дугою; ІХ, Х, ХІ пари черепних нервів (група блукаючого нерва) – з ІІІ, IV, V зябровими дугами.
ХІІ пара черепних нервів розвивається шляхом з’єднання нервових волокон верхніх шийних спинномозкових нервів і пов’язана з постбранхіальними (зазябровими або завушними) міотомами.
Клініцисти поділяють справжні черепні нерви на:
1) окорухові (III, IV, VI пари);
2) нерви мосто-мозочкового кута (V, VII, VIII пари);
3) каудальні нерви (ІХ, Х, ХІ, ХІІ пари).
Периферійні нерви утворені пучками нервових волокон, які об’єднані сполучнотканинними компонентами (оболонками): ендоневрієм, периневрієм і епіневрієм. Ендоневрій – це пухка сполучна тканина між окремими нервовими волокнами. У периневрії виділяють зовнішню частину – щільну сполучну тканину, яка оточує кожний пучок нервових волокон, і внутрішню частину – кілька концентричних шарів плоских периневральних клітин, ззовні і всередині вкритих товстою базальною мембраною, що містить колаген IV типу, ламінін, нідоген і фібронектин. Внутрішня частина периневрія (епітеліоїдний пласт периневральних клітин, з’єднаних за допомогою щільних контактів) утворює периневральний бар’єр, необхідний для підтримання гомеостазу в ендоневрії. Бар’єр контролює транспорт молекул через периневрій до нервових волокон, перешкоджає доступу в ендоневрій інфекційних агентів, захищає нервові волокна від пошкоджень при розтягненні нерва. У термінальній частині нерва периневрій обривається і має вигляд відкритої манжетки. Існує думка, що нейротропні віруси здатні проникати через цю ділянку у периневрій, розповсюджуючись по ньому в ЦНС.
Епіневрій – це зовнішня оболонка нерва, що зв’язує в ціле пучки нервових волокон (кількість яких залежить від діаметра нерва і може складати від одного до кількох десятків).
Розрізняють два види нервових волокон – безмієлінові і мієлінові. Обидва види волокон складаються з центрально розміщеного відростка нейрона (осьового циліндра), оточеного оболонкою із шваннівських клітин (лемоцитів). Клітини Шванна синтезують білки (Р0; Р1; Р2), утворюють мієлін і розгл ядаються як ана логи олігодендроцитів у ЦНС. При цьому, на відміну від олігодендроцита, кожна клітина Шванна мієлінізує один аксон. Мієлін у ПНС – це компактна структура зі змінених п ла змати чних мембран шваннівської к літини (в ЦНС – олігодендроцита), спірально закручених навколо аксонів. 80 % маси мієліну складають ліпіди, 20 % – білки мієліну: РО, Р22; основний білок мієліну; протеоліпідний тощо.
Безмієлінові нервові волокна в периферійному нерві у дорослого розміщуються переважно у складі вегетативної нервової системи і характеризуються відносно низькою швидкістю проведення нервових імпульсів (0,5–2 м/сек). Вони утворюються шляхом занурення осьового циліндра (аксона) у цитоплазму шваннівських клітин, розміщених у вигляді тяжів. При цьому плазмолема шваннівської клітини прогинається, оточуючи аксон, і утворює дублікатуру – мезаксон. Поверхня безмієлінового волокна вкрита базальною мембраною.
Мієлінові нервові волокна в периферійному нерві характеризуються високою швидкістю проведення нервових імпульсів (до 120 м/сек). Мієлінові волокна зазвичай товстіші за безмієлінові і містять осьові циліндри великого діаметра. В мієліновому волокні осьовий циліндр оточений мієліновою оболонкою, навколо якої розташована нейролема, утворена тонким шаром цитоплазми і ядром шваннівської клітини. Під електронним мікроскопом видно, що мієлінова оболонка виникає в результаті злиття численних (до 300) витків плазматичної мембрани шваннівської клітини навколо частини аксона.
Утворення мієлінової оболонки в периферійному нерві починається з занурення осьового циліндра у шваннівську клітину і супроводжується формуванням довгого мезаксона, який починає обертатись навколо аксона, утворюючи перші пухко розташовані витки мієлінової оболонки . По мірі збільшення числа витків у процесі дозрівання мієліну вони розташовуються щільніше і частково зливаються; проміжки між ними, заповнені цитоплазмою шваннівської клітини, зберігаються лише в окремих ділянках
– насічках Шмідта – Лантермана.
Сегменти мієліну відокремлені один від одного малими сегментами, в яких оголений аксон оточений інтерстиціальним простором. Ці сегменти, які називають вузлами (перехватами, або перетяжками) Ранв’є, є місцем знаходження множинних натрієвих каналів. Коли мембрана аксона збуджена, згенерований електричний імпульс не може проходити через високорезистентну оболонку мієліну і, отже, виходить назовні і деполяризує аксональну мембрану на наступному вузлі, який може мати протяжність 1 мм і більше. Тому розповсюдження деполяризації в мієліновому волокні здійснюється стрибками від вузла до вузла (сальтаторно). При цьому швидкість проведення збудження по мієліновому волокну прямо пропорційна і діаметру волокна, і довжині між вузлами Ранв’є (тобто чим більший діаметр і чим довші інтервали між вузлами Ранв’є, тим вища швидкість проведення збудження).
Нервове волокно характеризується збудливістю і лабільністю. Збудливість мієлінових волокон вища, ніж безмієлінових. Крім того, у мієлінових волокон вища лабільність порівняно з усіма іншими нервовими утвореннями, в тому числі і безмієліновими волокнами. Наприклад, відомо, що мієлінові волокна можуть відтворювати до 1000 імпульсів за 1 сек.
Виділяють такі закони проведення збудження по нервових волокнах:
1) закон ізольованого проведення збудження;
2) закон анатомічної і фізіологічної цілісності нервового волокна;
3) закон двостороннього проведення збудження;
4) закон практичної невтомлюваності нервових волокон;
5) закон прямо пропорційної залежності швидкості проведення імпульсу від діаметра нервового волокна.
Закон анатомічної і фізіологічної цілісності нервового волокна стверджує, що необхідною умовою проведення збудження у нерві є не тільки його анатомічна безперервність, але й фізіологічна цілісність. У будь-якому металевому провіднику електричний струм буде протікати доти, поки провідник зберігає фізичну безперервність. Для нервового “провідника” цієї умови недостатньо – нервове волокно повинне зберігати також фізіологічну цілісність. Якщо порушити властивості мембрани волокна (перев’язка, блокада новокаїном, аміаком тощо), то проведення збудження по волокну припиняється. Застосування в клінічній медицині з метою знеболювання місцевих анестетиків, які блокують активність натрієвих каналів, насамперед у вузлах Ранв’є, доводить, з одного боку, важливість даного закону, а з іншого – можливість зворотної блокади іонних каналів.
Закон двостороннього проведення збудження по нервовому волокну стверджує, що будь-яке нервове волокно (аферентне чи еферентне) здатне проводити збудження в обох напрямах (до нейрона чи від нього). У цьому можна переконатись, якщо штучно подразнювати волокна – потенціали дії будуть розповсюджуватися в обидва боки від місця подразнення. Однак реально за рахунок наявності одностороннього проведення збудження в хімічних синапсах всі нервові волокна проводять збудження в одному напрямку, характерному для даного волокна (по аферентних волокнах – у ЦНС, по еферентних волокнах – від ЦНС до органа).
Закон практичної невтомлюваності нервових волокон, сформульований Н. Є. Введенським, вказує на те, що нервове волокно володіє малою втомлюваністю. Дійсно, проведення імпульсу по нервовому волокну не порушується протягом тривалого (багатогодинного) експерименту. Вважають, що нервове волокно відносно невтомлюване внаслідок того, що процеси ресинтезу енергії в ньому ідуть з достатньо великою швидкістю і встигають відновити втрати енергії, що виникають при проходженні збудження.
У момент збудження енергія нервового волокна витрачається на роботу натрій-калієвого насоса. Особливо великі втрати енергії виникають у вузлах Ранв’є внаслідок великої щільності тут натрій-калієвих насосів.
Закон прямо пропорційної залежності швидкості проведення імпульсу від діаметра нервового волокна був встановлений лауреатами Нобелівської премії (1944) американськими фізіологами Джозефом Ерлангером і Гербертом Гассером. На основі цього закону автори запропонували широковідому класифікацію нервових волокон.