Структурні основи шкірної та суглобово-м’язової чутливостей
Шкіра (cutis) належить до загального покриву (integumentum commune), що безпосередньо стикається із зовнішнім середовищем і є надзвичайно важливим органом тіла, який бере різноманітну і активну участь у життєдіяльності організму.
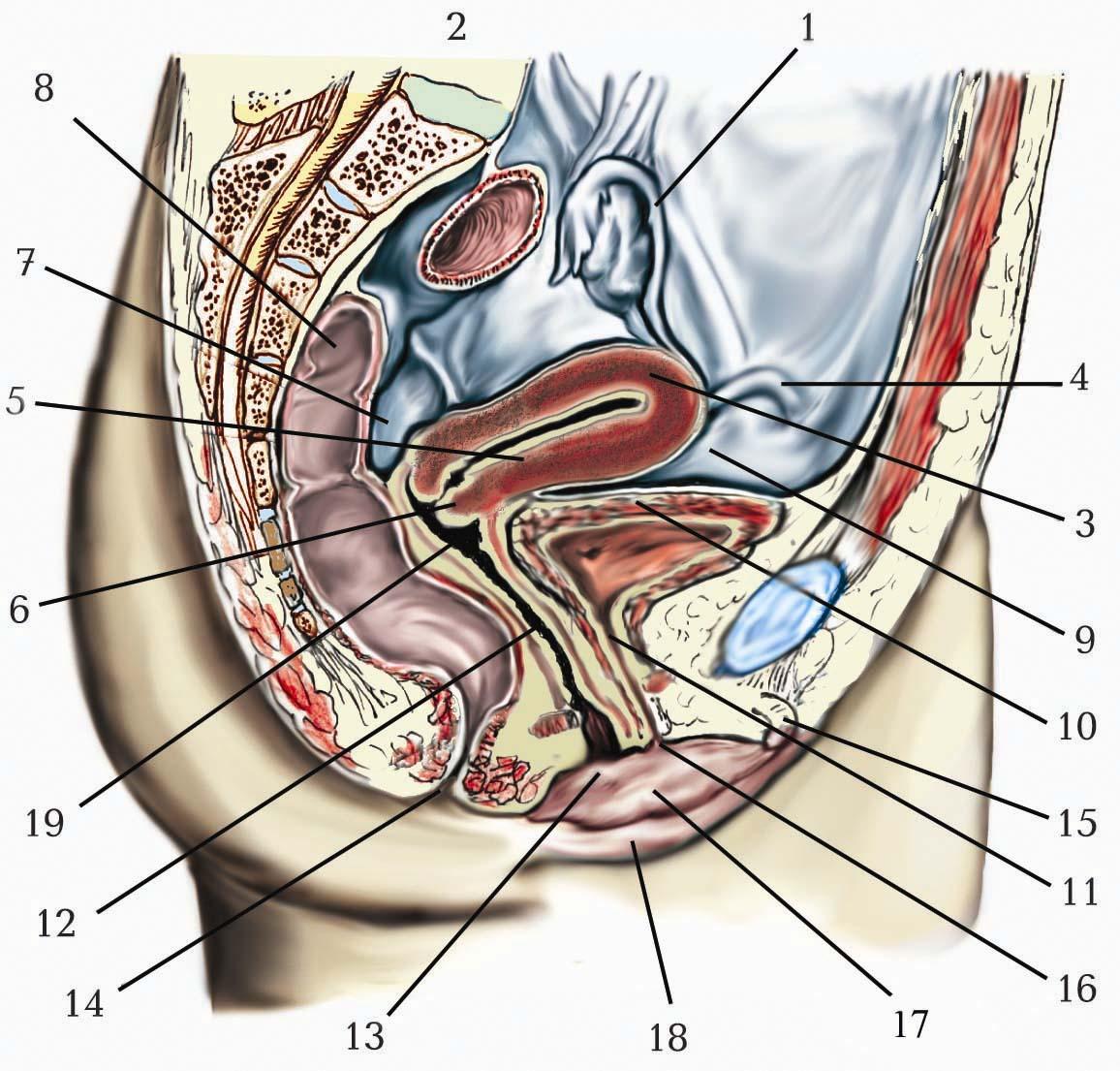
Нині загальнопідтверджено, що шкіра – це величезна поверхня рецепції з багатою і різноманітною чутливою іннервацією. Щодо іннервації шкіру ділять на окремі ділянки, межі яких не чітко відокремлені, а заходять і на сусідні території; крім того, нерви останніх зв’язані рядами анастомозів. Центри шкірних нервів, закладені у спинному і головному мозку, пов’язані з руховим апаратом, а також із внутрішніми органами. Кожний внутрішній орган має на шкірі свою приблизну зону (зону Захар’їна – Геда.) У цих зонах при захворюванні внутрішніх органів часто з’являються відображені болі, а також больова і температурна гіперестезія (підвищена чутливість до подразників). Установлені співвідношення між внутрішніми органами і сегментами шкірної іннервації: легені – 3–4 шийні, а також 2–5 грудні сегменти; серце – 3–5 шийні, 1–8 грудні, переважно зліва, інколи з обох боків; стравохід – в основному 5, а також 6–8 грудні; молочна залоза – 4–5 грудні; шлунок, підшлункова залоза – 7–9 грудні, звичайно з обох боків; тонка і товста кишки – 9–12 грудні з обох боків або тільки зліва; печінка – 3–4 шийні, 8–10 грудні справа; жовчний міхур – переважно 8–9 грудні, а також 5–7 грудні; нирка – переважно 10 грудний, а також 11–12 грудні, 1 поперековий; сечовід – 11–12 грудні, 1 поперековий; яєчка – 10 грудний; придаток яєчка – 11–12 грудні; сечовий міхур – 11–12 грудні, 1 поперековий, а також 3–4 крижові; передміхурова залоза – 10–11 грудні, а також 1–3 і 5 крижові; яєчник – 10 грудний; маткова труба – 11–12 грудні та 1–4 крижові; тіло матки – 10 грудний, перший поперековий сегменти.
Приймаючи до уваги, що, як встановлено на даний час, шкіра є не просто комплексним захисним покривом людського тіла, а й органом дотику і величезною рефлексогенною зоною, слід погодитися із зауваженням І. П. Павлова про те, що “різні точки шкіри є проекцією точок мозку”.
Шкіра має багату чутливу іннервацію. Найбільш масивне нервове сплетення розташоване у підшкірному прошарку. Від нього відгалужуються численні нервові волокна до волосся та залоз шкіри, а також до нервового сплетення в сосочковому шарі дерми. Це сплетення подано переважно мієліновими волокнами, що утворюють чутливі нервові закінчення в усіх шарах шкіри.
За Лаврентьєвим розрізняють вільні та невільні нервові закінчення. Вільні нервові закінчення – термінальні розгалуження периферійного відростка дендрита чутливого нейрона. Невільні нервові закінчення, окрім термінальних розгалужень дендрита чутливого нейрона, містять спеціальні клітини (практично всі закінчення цього типу – механорецептори). Інкапсульовані механорецептори (інкапсульовані тільця) – невільні закінчення, що мають оформлену сполучнотканинну капсулу.
Вільні нервові закінчення – найбільш поширений у шкірі тип рецепторів. У дермі вони зосереджені переважно в сосочках; вільні нервові закінчення надшкір’я (епідермісу) розташовані в базальному та шипуватому шарах; в ділянці шкіри з високою тактильною чутливістю (наприклад, пальці рук) терміналі досягають зернистого шару. Вільні нервові закінчення у надшкір’ї утворюють механорецептори, терморецептори та рецептори больової (ноцицептивної) чутливості. Терморецептори поділяють на рецептори холодової (25–30 °C) та теплової (40–42 °C) чутливості. Їх розгалужені терміналі проходять між глибокими шарами надшкір’я.
Загалом процес сприймання подразнень у шкірі відбувається за допомогою дуже різноманітних форм нервових закінчень, які, проте, є типовими для окремих територій та шарів шкіри. У базальному шарі епідермісу локалізуються комплекси клітин Меркеля з нервовими терміналями (цей тип механорецепторів виявлений у шкірі долонь та ступнів). Під епідермісом (у сосочковому шарі дерми) присутні дотикові тільця Мейснера, найбільш численні у шкірі долонної та підошвової поверхні пальців рук, ніг, а також у шкірі губ, повік, зовнішніх статевих органів та сосків грудної залози. У дермі також знаходиться другий тип інкапсульованих рецепторів – тільця Руффіні (вони найчастіше зустрічаються в шкірі підошвової поверхні ступні). В ретикулярному шарі дерми розташовуються колбочки Краузе. Тільця Фатера – Пачіні – найбільші інкапсульовані рецептори, локалізуються глибоко у дермі та підшкірному прошарку (переважно в пальцях, зовнішніх статевих органах та в молочній залозі).
Дотикові клітини Меркеля – округлі або подовжені клітини, що локалізовані в надшкір’ї. Вони більші, ніж епітеліоцити, ядро вигягнуте і сегментоване, цитоплазма світліша, в ній відсутні характерні для кератиноцитів скупчення проміжних філаментів. Клітини Меркеля поєднуються з епітеліоцитами за допомогою десмосом та формують контакт з нервовими терміналями. Встановлено, що клітина Меркеля може нести позиційну інформацію для визначення місця кінцевої локалізації терміналі периферійного відростка чутливого нейрона в нейрогенезі або при регенерації нервів.
Чутливі тільця Руффіні (corpuscula sensoria) – крупні рецептори веретеноподібної форми, довжиною до 2 мм та діаметром близько 150 мкм. Серцевину рецептора (внутрішню цибулину) утворюють розгалужені нервові терміналі, що оточені пластинчастими клітинами, аналогами шваннівських. Булавоподібні нервові терміналі не вкриті пластинчастими клітинами і виділені базальною мембраною від так званого капсулярного простору, який розташований між капсулою і внутрішньою цибулиною. Це досить великий простір, що заповнений рідиною, має фібробласти, макрофаги і неорієнтовані колагенові волокна, які вплітаються у внутрішню цибулину. Капсула тільця складається з 4–5 шарів ущільнених клітин. Іншим прикладом інкапсульованого рецептора в шкірі є пластинчасте тільце Фатера – Пачіні (corpusculum lamellosum). Основними складовими цього овального тільця (розмір до 0,5 × 1,0 мм) є внутрішня цибулина, зовнішня капсула та термінальне нервове волокно. Адекватна механічна стимуляція тільця Фатера – Пачіні призводить до появи рецепторного потенціалу в термінальній частині нервового закінчення. Досягнувши критичної величини, рецепторний потенціал у нервовому вузлі Ранв’є викликає появу потенціалів дії. Зовнішня капсула рецептора – фільтр, що пропускає тільки динамічну складову механічної дії. Тип чутливих нервових закінчень, що розглядається, відноситься до фазних рецепторів, які швидко адаптуються, при цьому потрібно пам’ятати, що тільця Фатера – Пачіні, як і всі тканинні механорецептори, – первинночутливі рецептори. Це означає, що первинний трансформаційний процес відбувається у нервовій терміналі рецептора.
На різних ділянках шкіри тактильна чутливість розвинена неоднаково. Поріг подразнення найчутливіших ділянок становить 50 мг, а в найменш чутливих він досягає 10 г. Найвищу чутливість мають губи, ніс, язик, найнижчу – спина, живіт, підошви, ступні. Для дотику характерне також просторове відчуття. Воно полягає у здатності розрізняти, сприймати як роздільні дві одночасно подразнювані точки. Пороги такого просторового розрізнення на різних частинах поверхні людського тіла дуже відрізняються. Встановлена також функціональна тактильна асиметрія правої і лівої руки. Помічено, що правші не тільки швидше і точніше виконують роботу правою рукою, а й краще розпізнають предмети на дотик цією самою рукою. Причина, мабуть, полягає в набагато більшому досвіді правої кінцівки, тобто сенсорна асиметрія є наслідком рухової асиметрії.
Слід зазначити, що дотикова чутливість, з одного боку, є одним із найдавніших видів чутливості і дуже добре розвинена у багатьох тварин; з другого боку, вона відіграла помітну роль у формуванні людини. На даний час загальновизнано, що чуття дотику як спосіб контакту із зовнішнім світом та одержання інформації про нього без перебільшення відіграє виняткову роль, тому що у взаємодії з іншими видами чуття, і насамперед із зором, дотик став основою для формування у людини цілісних уявлень про навколишні предмети і розвитку здатності до трудової діяльності.
Температурну сенсорну систему звичайно розглядають як частину шкірного аналізатора, і для того є достатні підстави. По-перше, рецептори цієї аферентної системи розташовані в шкірі; по-друге, як показали численні дослідження, їх не можна відділити від рецепторів тактильних. Дуже поширена думка, що подразнення тілець Руффіні спричиняється до відчуття тепла, а колбочок Краузе – холоду. Разом з тим установлено, що до температурного впливу виявилися чутливими і ділянки шкіри, в яких містяться тільки вільні нервові закінчення.
Слід зауважити, що й холодові, і теплові рецептори адаптуються дуже слабо. Це означає, що за тривалої дії сталої температури зберігається незмінною частота імпульсів, які вони посилають. Звідси випливає дуже важливе для розуміння фізіології терморегуляції положення – теплові і холодові рецептори є датчиками абсолютної температури, а не її відносних змін. При цьому крайні і холодові, і теплові відчуття без різкої межі переходять у відчуття болю.
Біль характеризується дуже істотною своєрідністю, яка відрізняє його від відчуттів, пов’язаних з функціонуванням усіх раніше розглянутих органів чуття. Така своєрідність, по-перше, полягає у тому, що для болю немає специфічних за модальністю (видом енергії) адекватних подразників (це можуть бути й механічні, і теплові, і хімічні впливи). По-друге, больова чутливість притаманна багатьом органам і тканинам людини. В уявленнях про загальні механізми болю тривалий час співіснують дві протилежні точки зору. Одна з них – теорія специфічних больових шляхів – заснована на твердженні, що біль є самостійним відчуттям. Згідно з цією концепцією, остаточно сформованою у 1895 р. М. Фреєм, біль є результатом збудження спеціальних больових рецепторів і має як власні специфічні шляхи проведення больової імпульсації, так і специфічні нервові центри. Друга концепція, що була висунута у 1894 р. А. Гольдшейдером, стверджує неспецифічну природу механізму больового збудження і розглядає виникнення болю як результат сумації сенсорних подразнень, що викликають сильну дію і тим самим біль. Всі інші теорії, що виникли пізніше, практично являють собою різноманітні модифікації цих двох концепцій.
Особливо слід відзначити точку зору філософа та психолога Г. Маршалла, який у 1894 р. висловив свою думку таким чином: “Біль – емоційна якість, що забарвлює всі сенсорні явища”. Він визнавав, що існує колюче і ріжуче відчуття, але вважав, що біль суттєвим чином від них відрізняється. Всі сенсорні шляхи, а також думки можуть мати больову межу, і Г. Маршалл казав про біль втрати й про те, як боляче чути музику, що погано виконується. Такі крайні погляди, звісно, викликали критику. Ч. Шеррінгтон у 1900 р. вказував, наприклад, що біль обпеченої руки відрізняється від того “болю”, який відчуває меломан навіть при дуже жахливому дисонансі. Однак Ч. Шеррінгтон розумів, що у болю є як сенсорна, так і афективна межа, і виразив це так: “Мозок рідко сприймає (вірогідно, ніколи не сприймає) будь-що з повною байдужістю, тобто без “відчуття” … афективне забарвлення притаманне всім відчуттям, тому до властивостей шкірного відчуття належить і шкірний біль”.
Нині доведено, що більшість нервових утворень, які збуджуються ноцицептивними (від латинського nocere – пошкоджувати) стимулами, реагують і на подразнення, що не пошкоджують. У той же час неодноразово описувались окремі нейрони спинномозкових вузлів, що відповідають тільки на ноцицептивні стимули. Для центральної нервової системи такі властивості нейронів виявлені менш виразно і їх існування не визнається остаточно доведеним (Л. Н. Смолин, 1975). Для більшості цих нейронів у ЦНС характерна конвергенція як сенсорних, так і ноцицептивних імпульсацій, тому до останнього часу і концепція Фрея, і концепція Гольдшейдера мають право на існування.
Відомо, що всі соматичні рецептори за порогом чутливості поділяються на низькопорогові і високопорогові. Низькопорогові рецептори активно збуджуються подразниками, що не пошкоджують (дотик, тиск), а при посиленні подразнення до пошкоджуючого ступеня спостерігається навіть послаблення їх реакції. Однак і низькопорогові рецептори, що передають імпульс по А, альфаі А, бета-волокнах, також можуть бути ноцицепторами, оскільки при повторній стимуляції, що викликає залучення лише А, альфа-волокон, спостерігається виникнення больового відчуття у людини.
Високопорогові рецептори, як правило, збуджуються при дії сильних пошкоджуючих подразників (сильне стискання, уколи, розтини, сильні температурні дії, впливи хімічних речовин тощо), але можуть реагувати й на стимули, що не пошкоджують. Саме тому високопорогові рецептори отримали назву больових рецепторів або ноцицепторів. Вважають, що ноцицептори являють собою вільні нервові закінчення немієлінізованих волокон, що утворюють плексиморфні сплетення в шкірі. Знайдено й тонкі нервові волокна, що пов’язують їх з рецепторами дотику, тиску й температури (волокна Тимофєєва), що може служити основою збудження ноцицепторів при сильних сенсорних подразненнях.
Імпульси збудження від больових рецепторів проводяться до ЦНС двома видами нервових волокон. Вважають, що аферентна імпульсація, пов’язана з гострим локалізованим (дискретним) колючим болем, поширюється тонкими мієліновими волокнами типу А, а з пекучим дифузним тривалим болем – тонкими немієлінізованими волокнами типу С (швидкість проведення в останніх менша). У спинному мозку імпульси гострого локалізованого болю поширюються бічним спинномозково-таламічним шляхом (tractus spinothalamicus lateralis), а імпульси дифузного тривалого болю – спинномозковосітчастим шляхом (tractus spinoreticularis).
Незважаючи на різноманітність структур, що беруть участь у формуванні болю як стану, особливу роль відводять таламусу. Саме з цим утворенням пов’язана взаємодія між дифузною і дискретною системами формування відчуттів болю, регуляція аферентного потоку. Ось чому при порушенні цієї функції розвиваються дуже тяжкі, нестерпні «центральні» болі, які виникають без будь-яких зовнішніх впливів або при дуже незначних.
Проте в ЦНС є й так звані антиноцицептивні системи, до яких, зокрема, належить сітчаста формація (ядра шва) та утворення лімбічної системи, електричне подразнення яких в експерименті призводить до знеболювання. Механізмом такої антиноцицептивної дії є, мабуть, гальмування нейронів високопорогової дифузної аферентації.
Спроба поєднати структурні і функціональні аспекти альтернативних концепцій болю була здійснена канадським психологом і нейрофізіологом Р. Мелзаком і англійським нейрофізіологом П. Уоллом, які у 1965 р. запропонували ворітну теорію болю. По суті ця теорія припускає (Р. Мелзак, 1981), що нейронний механізм, який знаходиться в задніх рогах спинного мозку, – драглиста речовина (substantia gelatinosa) – діє як ворота, які можуть збільшувати або зменшувати потік нервових імпульсів, що йдуть від периферійних волокон в ЦНС. Таким чином, соматичний вхід піддається моделюючому впливу воріт до того, як він викличе сприйняття болю і відповідну реакцію. В якому ступені ворота збільшують чи зменшують передачу імпульсів, визначається співвідношенням активності волокон діаметра А, бета і малого діаметра (А, дельта і С), а також низхідним впливом головного мозку. Коли кількість інформації, що пройшла через ворота, перебільшує критичний рівень, збуджуються нейронні ділянки, що беруть участь у відчутті болю і у відповідній реакції на нього.
Важливим висновком ворітної теорії стосовно шкіри є визнання того, що біль є негативною біологічною потребою, що формує перцептуально-мотиваційний компонент тієї функціональної системи, яка контролює цілісність шкіри як покривної оболонки організму, що забезпечує ізольованість його від зовнішнього світу і тим самим зберігає сталість внутрішнього середовища організму.
Анатомічні та фізіологічні дані свідчать про те, що система шкірної чутливості – це єдина інтегрована система, що складається із спеціалізованих складових частин. Декілька паралельних систем одночасно аналізують вхід, щоб забезпечити різноманітність відтінків та складність відчуттів, що зазнаються, а також відповідної реакції. Деякі ділянки (шари) шкіри володіють особливою спеціалізацією, що дозволяє вибирати сенсорно-дискримінаційну інформацію, тоді як інші ділянки грають особливу роль в мотиваційноафективних проявах (біль). Ці паралельні системи обробки інформації взаємодіють одна з одною та повинні взаємодіяти також з тими видами кіркової активності, що пов’язані з минулим життєвим досвідом, увагою та іншими когнітивними процесами. Ці процеси, що взаємодіють, створюють те незліченне число патернів активності (патерн у буквальному перекладі – узор, і у фізіології патерн означає просторово-часовий розподіл імпульсивної активності), які служать основою для різноманітних шкірних відчуттів.
Життя людини неможливе без рухів: так, коли людина спокійно стоїть, сидить і навіть лежить, її скелетні м’язи не бувають у стані цілковитого спокою. Регулятором рухів і водночас чуттям, яке допомагає людині узнавати в кожний даний момент положення тіла та його частин у просторі, є суглобово-м’язове (пропріоцептивне) чуття. Специфічними рецепторами (пропріорецепторами) опорно-рухового апарату є нервово-сухожилкове веретено (сухожилковий орган Гольджі) та нервово-м’язове веретено (м’язове веретено).
Нервово-сухожилкові веретена розташовуються в сухожилках звичайно на межі м’язової тканини, в суглобових капсулах та зв’язках і реагують на виникнення розтягнення в них. Нервово-сухожилкове веретено ніколи не буває у стані спокою і постійно відображає стан опорно-рухового апарату, але ступінь його збудження буде тим сильніший, чим інтенсивніше скорочення м’яза. Таким чином, для будь-якого певного положення суглобів існує своя специфічна картина аферентної імпульсації, яка надходить від нервово-сухожилкового веретена до кори півкуль мозочка та великого мозку по пропріоцептивних шляхах. На відміну від нервово-сухожилкового веретена, розташованого “послідовно” між м’язом і сухожилком, нервово-м’язове веретено в цьому ланцюгу розміщене “паралельно”. Цілком зрозуміло, що нервово-м’язове веретено буде розтягуватися і збуджуватися під час розслаблення м’яза, а ступінь його збудження буде пропорційна ступеню розслаблення м’яза. Наявність таких двох різних пропріорецепторів дає можливість одержувати тонко диференційовану інформацію про стан м’яза, тобто ступінь його скорочення, розслаблення або розтягнення. Коли м’яз розслаблений, відбувається слабка тонічна аферентна імпульсація від нервово-сухожилкових веретен і посилена – від нервово-м’язових веретен (при скороченні м’яза встановлюється протилежне співвідношення, а при штучному розтягненні аферентація посилюється від обох видів пропріорецепторів).
Кожне нервово-м’язове веретено складається, як правило, з кількох (1–10) так званих інтрафузальних м’язових волокон. Існує два типи інтрафузальних м’язових волокон: ЯС-волокна, у яких ядра сконцентровані в центральній частині у вигляді ядерної сумки, і ЯЛ-волокна з розташуванням ядер у вигляді ядерного ланцюжка .
Інтрафузальні м’язові волокна одержують як чутливу, так і рухову (від гамма-мотонейронів) іннервацію. Функція гамма-мотонейронів полягає у пристосуванні скорочення м’язових волокон у веретені до стану скелетного м’яза. Імпульсація гамма-еферентних нейронів, як правило, знижує поріг збудження нервово-м’язових веретен. Цей механізм відіграє важливу роль у підтриманні певного положення тіла.
Гамма-еферентний механізм складає частину саморегульованої системи. Коли тіло або яка-небудь частина його приймає певне положення, розтягненню відповідних м’язів передує прихід гамма-еферентної імпульсації, яка регулює частоту імпульсів, що породжуються нервово-м’язовими веретенами. Потім центрально керовані моторні нерви викликають скорочення відповідних м’язів, причому активність цих нервів контролюється рефлекторно гамма-еферентами. Таким чином, ця система (її називають також гамма-петлею) містить один аферентний і два еферентних шляхи.
Що стосується підтримки певного положення тіла, то як контрактильний (від лат. contractio – стискання, скорочення) м’язовий тонус, так і активні скорочення м’язів здійснюються лише за наявності чіткого узгодження в діяльності сегментарного апарату спинного мозку. Воно виражається у реципрокній (спільній, протилежно спрямованій) іннервації м’язів, що забезпечують рух та узгоджене скорочення м’язів для фіксації суглобів. Так, наприклад, при згинанні одночасно з напруженням згиначів відбувається реципрокне розслаблення тонусу антагоністів-розгиначів і, навпаки, при розгинанні – реципрокне розслаблення згиначів. При виконанні дрібних рухів кистю одночасно сама собою фіксується рука у ліктьовому та плечовому суглобах шляхом узгодженого скорочення відповідних м’язів. Можна привести безліч подібних прикладів, адже в кожному русі людини ці механізми є.
Кожному руховому акту, в тому числі й довільному, передує позиційне збудження та установлення м’язів, що дає можливість виконати рух чітко і без шкоди для організму. Підніманню, наприклад, лівої ноги передує напруження м’язів правої ноги і тулуба для утримання рівноваги. Це все здійснюється автоматично, без участі свідомості, однак лише доти гладко та бездоганно, поки збережені всі нервові зв’язки, аферентні та еферентні нейрони.
Адаптація у руховому аналізаторі на рецепторному рівні виявлена слабо. Завдяки цьому аферентна імпульсація тривалий час не змінюється при незмінному ступені розтягнення пропріорецепторів. Протеінтегральна чутливість сенсорної системи в цілому змінюється залежно від навантаження на опорно-руховий апарат. Відома його здатність до тренування, що виявляється в розвиткові тонкої рухової координації відповідних груп м’язів у музикантів, хірургів тощо.
Загальний покрив
Загальний покрив (integumentum commune) тіла людини утворений шкірою, в якій закладені потові та сальні залози, чутливі нервові закінчення, волосся та нігті.
Шкіра
Шкіра (cutis) виконує захисну функцію, бере участь у терморегуляції та виділенні шкідливих речовин з потом, обміні речовин та газообміні, накопиченні енергетичних запасів. Займаючи велику площу (в середньому близько 1,8 м2), шкіра сприймає різні подразнення, головними з яких треба вважати тактильні, больові й температурні; але, без сумніву, на шкіру впливають подразники світлові й звукові. Шкіра складається з поверхневого шару – надшкір’я та глибокого шару – дерми, або власне шкіри. Підшкірний прошарок, або підшкір’я, складається з підшкірної жирової клітковини та пухкої сполучної тканини. Товщина самої шкіри коливається в межах від 0,5 мм до 0,5 см. Найтовстіша шкіра – на сідницях, підошвовій поверхні стопи, потилиці; найтонша – шкіра cтатевого члена, шкіра повік. Маса шкіри дорослої людини, загалом беручи, варіює в межах приблизно 3000–5000 г, тоді як підшкірний прошарок варіює ще більше, і вага його у дорослої людини становить 12–20 кг (наведені цифри приблизно визначають середні розміри).
Надшкір’я (epidermis) представлене багатошаровим плоским зроговілим епітелієм, завтовшки від 30 мкм до 4 мм. У надшкір’ї розрізняють п’ять шарів (із зовнішньої поверхні вглиб): роговий, блискучий, зернистий, шипуватий та базальний.
Поверхневий роговий шар (stratum corneum) складається з плоских зроговілих клітин, які на поверхні надшкір’я постійно злущуються і відпадають.
Глибший блискучий шар (stratum lucidum) складається з 3–4 рядів клітин, що заповнені блискучою речовиною – елеїдином. При затвердінні елеїдину формується кератин, ядро клітини зникає, у ній припиняється обмін речовин, і вона перетворюється на лусочку рогового шару.
Третій зернистий шар (stratum granulosum) складається з 1–5 рядів клітин з дегенеративними змінами у ядрах та кератогіаліновими зернятками (granulum keratohyalini) у цитоплазмі.
Шипуватий шар (stratum spinosum) складається з шипуватих епідермоцитів (epidermocytus spinosus), що з’єднані між собою за допомогою шипуватих відростків.
Найглибший базальний шар (stratum basale) cкладається з 5–15 рядів росткових клітин, за рахунок яких відбувається поновлення вищерозміщених шарів надшкір’я. Крім основних епідермоцитів, цей шар містить також меланоцити, кількість пігменту в яких обумовлює колір шкіри.
Дерма (власне шкіра), derma (corium) побудована із пухкої волокнистої сполучної тканини, в якій містяться колагенові, еластичні та непосмуговані м’язові волокна. Від кількості еластичних волокон у дермі залежить еластичність шкіри. Дерма пронизана великою кількістю судин та нервів. Розрізняють поверхневий (сосочковий) шар та глибокий (сітчастий) шар дерми.
Сосочковий шар (stratum papillare) має виступи – сосочки (papillae), які зв’язують цей шар дерми з надшкір’ям. У сосочках розміщені кровоносні капіляри, нервові закінчення та сліпі вирости лімфокапілярів. Ряди сосочків виступають над поверхнею шкіри, утворюючи гребінці шкіри (cristae cutis), на верхівці яких відкриваються протоки потових залоз. Гребінці шкіри формують генетично обумовлений, типовий для кожної людини малюнок на поверхні кінчиків пальців, долонь та підошов. Цей малюнок (папілярні лінії) становить основу для багатьох досліджень у галузі медичної генетики та в криміналістиці (дерматогліфіка, дактилоскопія).
Сітчастий шар (stratum reticulare), що побудований з неоформленої щільної сполучної тканини і складається з сітки щільно переплетених колагенових волокон та
невеликої кількості еластичних і ретикулярних волокон, без чіткої межі продовжується у підшкірний прошарок.
Підшкірний прошарок (tela subcutanea), або підшкір’я (hypodermis) містить підшкірну жирову клітковину (panniculus adiposus) та пухку сполучну тканину (textus connectivus laxus). Жирові відкладення підшкірного прошарку виконують функцію термоізолятора та депо енергетичних запасів.
У підшкірному прошарку описують також м’язовий шар (stratum musculorum), волокнистий шар (stratum fibrosum) та перетинчастий шар (stratum membranosum). Жоден з компонентів підшкірного прошарку не є наявним на всіх ділянках тіла. Наприклад, у калитці підшкірний прошарок представлений тільки м’язовим шаром, який утворює її м’ясисту оболонку (tunica dartos).
Найбільш широко розповсюдженою є підшкірна жирова клітковина, яка є пластичною масою, що формує обриси тіла. Насамперед треба відзначити, що існують ділянки шкіри, де жир завжди відсутній, і, навпаки, є місця, схильні до надмірного жировідкладання. Наприклад, вушна раковина, повіка, грудний сосок, шкіра статевого члена, навіть у найугодованіших суб’єктів, завжди не мають жиру. Ділянка щоки, підщелепна (під підборіддям), ділянка грудної залози, передня черевна стінка, сіднична ділянка і передня стегнова ділянка є тими ділянками, де жир завжди існує до моменту крайнього виснаження людини і де у жирних людей кількість його може досягати неймовірних розмірів.
У дуже вгодованих людей сильне відкладання підшкірного жиру спостерігається під підборіддям. Шкіра над ним збирається в одну або дві поперечні складки і утворює так зване подвійне або потрійне підборіддя. Неоднакова підшкірна жирова тканина в різних відділах грудної стінки. Лежачи досить тонким пухким шаром у задньому і бічному відділі, у жінок вона спереду потовщується і, роздвоюючись на два шари, охоплює спереду і ззаду грудну залозу, причому жир зазвичай буває найбільше розвинений по обводу основи залози. В напрямі до верхівки залози кількість жиру зменшується, і навколо соска і на соску жиру немає. Іноді майже вся маса грудної залози проростає жировою тканиною; при цих умовах жінка, незважаючи на сильно розвинені пружні груди, мало здатна годувати дитину, тоді як інші, в яких груди значно менші, можуть бути добрими годувальницями.
Кількість жиру дуже неоднакова під шкірою передньої черевної стінки. Тут, у дуже легко розтяжній пухкій підшкірній клітковині, скупчення жиру звичайно пропорційне ступеню загальної вгодованості.
Особливістю підшкірного жиру промежини є безпосереднє сполучення з жировим скупченням у глибині промежини, біля прямої кишки, у так званій fossa ischioanalis. Із цим жировим скупченням сполучається жирова клітковина, що оточує всі органи малого таза. Звідси зрозумілі шляхи, якими гній при запаленні цих органів проривається під шкіру промежини.
У значній мірі розвинений підшкірний жир у всіх людей у сідничній ділянці; він утворює ніби товсту пружну подушку, яка у деяких народностей доходить до дуже великих розмірів.
Ті місця, які весь час перебувають під тиском (долоня, підошва, сіднична частина), є місцями особливого спеціального (коміркового) структурного диференціювання жирової маси. Коміркова структура жирового шару найкраще захищає тканини і органи, які лежать під ним, від повсякчасного тиску: міцні фіброзні пучки, які становлять стінки комірок, спресовують вміщені в них жирові грудки і створюють з жиру пружну еластичну подушку. Коли розподілити місця, які підпадають тиску, за ступенем його сили й тривалості, то першою треба поставити підошву, потім долоню і, нарешті, сідничну ділянку. Ступінь виявленості коміркової структури жиру в цих місцях прямо пропорційний частоті і силі механічного навантаження на вказані ділянки.
Шкіру з підлеглими тканинами (фасцією або окістям кісток) зв’язують пучки сполучнотканинних волокон – утримувачі шкіри (retinacula cutis). Один із добре розвинених утримувачів – хвостовий утримувач (retinaculum caudale) – йде від куприка до шкіри над ним, утворюючи на шкірі неглибоку куприкову ямочку (foveola coccygea). Хвостовий утримувач є рудиментарною структурою у людини, він розвивається із каудального кінця ембріональної хорди.
Придатки шкіри (волосся та нігті) та залози шкіри є похідними надшкір’я.
Волоссям (pili) різною мірою вкрита шкіра всього тіла людини. Волосяний покрив є розвиненим у багатьох ссавців (шерсть) і виконує у них теплоізолюючу функцію. У людини розрізняють такі види волосся: пушок (lanugo), брови (supercilia), вії (cilia), борода (barba), волосся вуха (tragi), волосся ніздрів (vibrissae), пахвове волосся (hirci), лобкове волосся (pubes).
У волосині розрізняють корінь (radix pili), розміщений у товщі шкіри, та стрижень (scapus pili), який виступає над поверхнею шкіри. Корінь волосини (корінь волоса) оточений сполучнотканинним волосяним мішечком (folliculus pili), у який відкривається сальна залоза. Приблизно до середини волосяного мішечка прикріплюється косий пучок непосмугованих м’язових волокон, що бере початок від сосочкового шару дерми, – м’яз-випрямляч волоса (m. arrector pili). При скороченні цього м’яза стрижень волосся підіймається над шкірою і у людини утворюється “гусяча шкіра”. Стискання сальної залози під час випрямлення волоса сприяє її змащенню сальним секретом. Колір волосся залежить від кількості у ньому пігменту меланіну та повітря. Збільшення кількості повітря та зменшення кількості меланіну у волоссі призводить до його посивіння.
Ніготь (unguis) є похідним надшкір’я і розвивається через зроговіння його клітин. Ніготь лежить на ложі нігтя (matrix unguis), яке сформоване глибокими шарами надшкір’я та дермою. Ріст нігтя відбувається в його корені (radix unguis), який складається з незроговілих ще клітин; ця ділянка має форму півмісяця і відрізняється білуватим кольором, у зв’язку з чим дістала назву місячка (lunula). Зроговіла частина нігтя має рожевуватий колір, тому що крізь товщу рогу просвічують кровоносні капіляри нігтьового ложа.
Залози шкіри (glandulae cutis) ділять на потові залози (glandulae sudorifera) та сальні залози (glandulaе sebacea). Спостерігаються ділянки шкіри, де переважають потові залози (які взагалі відсутні в червоній обвідці губ, головці та передній шкірочці статевого члена), і інші ділянки, з переважанням сальних залоз (які взагалі відсутні на підошвах та долонях). Загальновизнано, що потові залози, виділяючи піт, у значній мірі полегшують роботу нирок, а сальні залози змащують шкіру, захищаючи її від висихання.
Розвиток і аномалії розвитку шкіри
Шкіра має подвійне походження: поверхневий шар (надшкір’я) розвивається з поверхневої ектодерми; глибокий шар (дерма) розвивається з прилеглої мезенхіми. Протягом перших трьох місяців пренатального розвитку в надшкір’я проникають клітини, які походять з нервового гребеня. Ці клітини синтезують пігмент меланін, який може переноситися в інші клітини надшкір’я по розгалужених відростках. Після народження ці клітини, що мають назву меланоцитів, викликають пігментацію шкіри.
При народженні шкіра вкрита білуватою сироподібною змазкою, яка складається з секрету сальних залоз, відмерлих клітин надшкір’я та волосся. Сироподібна маса захищає шкіру від мацеруючої дії навколоплідних вод.
До аномалії розвитку шкіри належить іхтіоз – надмірна кератинізація шкіри, характерна для групи спадкових розладів, що зазвичай передаються як аутосомно-рецесивна ознака, проте можуть бути пов’язані і з X-хромосомою. У важких випадках ця патологія може призводити до гротескного вигляду новонародженого, як, наприклад, плід-арлекін. У плода-арлекіна шкіра покрита товстими роговими щитками, що розділені борознами та тріщинами.
Волосся з’являється на четвертому місяці пренатального розвитку як щільні вростання надшкір’я вглиб прилеглої дерми. Первинне пушкове волосся, яке має назву пушка (lanugo), випадає до або одразу після народження і пізніше заміщується грубішим пушковим волоссям, яке росте з нових волосяних мішечків.
До аномалій розвитку волосся належить гіпертрихоз (від грецького trichos – волосся) – збільшений волосяний покрив, зумовлений надлишковою кількістю волосяних мішечків. Він може локалізуватися на певних ділянках тіла (особливо у поперековій ділянці, вкриваючи приховане розщеплення хребта) або мати загальне розповсюдження, вкриваючи усю поверхню тіла.
Атрихія – вроджена відсутність волосся, як правило, пов’язана з аномаліями інших похідних ектодерми, таких як зуби та нігті.
Грудь. Грудна залоза
Грудь (mamma) функціонально тісно зв’язана зі статевою системою жінки і призначена для вигодовування жінкою новонародженого. В сучасній анатомічній номенклатурі термін чоловіча грудь (mamma masculina) виключений, тому що груди у чоловіків містять ті ж самі елементи, що й у жінок, тільки недорозвинені.
Грудь статеводозрілої (що не народжувала) жінки має вигляд більш або менш правильної півкулі, розміщеної на передній поверхні m. pectoralis major і частково на m. serratus anterior між III і VI (рідше VII) ребром. Грудь лежить на грудній фасції, що вкриває великий грудний м’яз, і з’єднана з цією фасцією тонким шаром пухкої сполучної тканини (тому є достатньо рухомою). Сполучнотканинні волокнисті тяжі, що йдуть від шкіри груді, носять назву підвішувальних зв’язок груді або тримача шкіри груді (ligg. suspensoria mammaria seu retinaculum cutis mammae). Груди займають значний простір між грудниною і пахвою, причому між правою і лівою спереду груднини лишається міжгрудна борозна (sulcus intermammarius). Маса груді незайманої 150–200 г, матері-годувальниці – вдвоє більша. Шкіра грудей ніжна, тонка (через неї легко просвічують вени), рухома. Приблизно на середині груді, на рівні IV міжреберного проміжка або V ребра, знаходиться грудний сосок (papilla mammae), переважно конічної форми: він лежить у центрі пігментованої ділянки шкіри – грудного кружальця (areola mammae). Добре розвинутою груддю вважається та, що невелика, пружна, тримається прямо, не звисаючи донизу, сосок спрямований вперед і трохи вбік. Величина і форма грудей мають не тільки вікові та індивідуальні відмінності, але змінюються від положення тіла (наприклад, у лежачому положенні грудь помітно стає більш плоскою), від ступеня вгодованості суб’єкта (навіть у дівчини, якщо вона дуже виснажена, груди зменшуються в розмірах і відвисають). Дуже часто спостерігається асиметрія грудей.
Тіло груді (corpus mammae) складається з жирової тканини та залозистих елементів, які формують грудну (молочну) залозу (glandula mammaria). Розміри грудної залози не відповідають величині груді: насправді грудна залоза далеко менша, бо оточена з усіх боків жировою клітковиною. При патології жировідкладення та при ожирінні груди через скупчення жиру можуть досягати 12 і більше кілограмів ваги.
Грудна залоза має пахвовий або бічний відросток (processus axillaries seu processus lateralis), який досягає пахвової ямки. Грудна залоза складається з 15–20 конусоподібних часток грудної залози (lobi glandulae mammariae), верхівки яких обернені до соска. Частки залози за будовою належать до складних альвеолярно-трубчастих залоз, вони розділені прошарками пухкої сполучної та жирової тканин. Частки залози та перегородки між ними спрямовані по відношенню до соска радіально. Частки побудовані з менших часточок грудної залози (lobuli glandulae mammariae), що розділені тонкими сполучнотканинними прошарками. Вивідні протоки часток грудної залози називаються молочними протоками (ductus lactiferi). Є 15–20 молочних проток, які мають діаметр 1,7–2,3 мм і відкриваються на соску. Поблизу місця відкриття на соску молочні протоки розширюються до 5–8 мм в діаметрі, утворюючи молочні пазухи (sinus lactiferi). По поверхні грудного кружальця розкидані невеликі горбки, на верхівці яких відкриваються протоки потових кружальцевих залоз (glandulae areolares).
Сосок і грудне кружальце вкриті продовженням шкіри груді; дерма утворює численні високі (особливо на соску) сосочки, які містять петлі кровоносних капілярів і закінчення чутливих нервів. Крім сполучної тканини, до складу соска та грудного кружальця входить гладка м’язова тканина, пучки якої почасти розміщені циркулярно навколо кінців ductus lactiferi, частково піднімаються прямовисно у товщі соска: внаслідок цього останній може рефлекторно змінювати свою форму і консистенцію – твердне.
Функціонуючу грудну залозу можна спостерігати тільки при вагітності. Груди починають змінюватись уже на другому місяці вагітності, на четвертомуп’ятому це стає особливо помітним, грудне кружальце і сосок темніють, glandulae areolares видно ясніше, груди збільшуються, їх шкіра напружується. Кровоносні і лімфатичні судини розширюються, сполучна тканина спочатку також зростає, але потім посилено проліферує залозиста частина грудей: ступінь розгалуження молочних проток збільшується, з’являються нові паростки. Період годування триває різний час, після чого настає зворотний розвиток залозистої речовини; при цьому значна частина кінцевих відділів атрофується. Величина груді зменшується, але початкова форма її (дівоча грудь) уже не може відновитися, навіть якщо жінка, народивши дитину, сама її не годувала.
Розвиток, вікові особливості й аномалії розвитку груді
Перші ознаки появи грудних залоз спостерігається у вигляді поздовжнього потовщення епідермісу – так званої молочної лінії, або молочного гребеня. У семитижневого зародка ця лінія тягнеться вздовж обох боків тулуба від основи верхньої кінцівки до ділянки відродження нижньої кінцівки. Хоча більша частина молочної лінії швидко зникає одразу після свого утворення, невелика частина її, розміщена у грудній ділянці, залишається та вростає у прилеглу мезенхіму. Тут вона формує від 16 до 24 тяжів, які, у свою чергу, дають початок невеликим щільним брунькам. Наприкінці пренатального періоду епітеліальні тяжі перетворюються на канали і формують молочні протоки, у той час як бруньки трансформуються на дрібні протоки та залозисті альвеоли. Спочатку молочні протоки відкриваються у невелику епітеліальну ямку. Невдовзі після народження дитини ця ямка перетворюється на сосок внаслідок проліферації прилеглої мезенхіми.
У новонародженого система вивідних проток розвинута добре, але кінцеві відділи часточок (секреторні) ледве намічені; сосок ще не сформований. У суб’єктів чоловічої статі сосок формується проліферація молочного гребеня пізніше, висота його у дорослого не перевищує 5 мм. У дівчаток в ділянці груді нагромаджується поступово жирова клітковина, що проростає між зачатками залозистих часточок. На момент статевої зрілості розвиток жирової тканини проходить особливо сильно, росте сосок і саме тіло груді, яка набуває типової зовнішньої форми і величини; проте грудь містить переважно фіброзну тканину і розгалужені вивідні протоки; залозисті часточки слабо розвинені. В такому бездіяльному стані грудь може лишатися довгі роки, і, якщо не настає вагітність, то з віком вона редукується: зменшується в розмірах її залозиста частина і навкружна жирова клітковина; у старих жінок груди мають вигляд двох в’ялих складок шкіри з соском і пігментованим віночком.
З аномалії розвитку грудей частіше зустрічаються політелія, полімастія, інвертований сосок.
Політелія – патологічний стан, для якого характерне утворення додаткових сосків внаслідок збереження фрагментів молочної лінії. Додаткові соски можуть виникати у будь-якому місці вздовж первинної молочної лінії, але зазвичай вони з’являються у пахвовій ділянці.
Полімастія має місце тоді, коли залишки молочної лінії перетворюються на сформовані грудні залози.
Інвертований сосок – це патологічний стан, при якому молочні протоки відкриваються у первісну епітеліальну ямку, що не перетворилася на повноцінний сосок.
У чоловіків іноді спостерігається надлишковий розвиток грудей (та грудних залоз) з обох або з одного боку. Груди при цій аномалії (яка дістала назву гінекомастії) нагадують формою і величиною груди жінок.