Залози внутрішньої секреції (glandulae endocrinae)
Ендокринологія (від грецьких слів: endon – усередині, krino – виділяти, logos – наука, вчення) – наука про залози внутрішньої секреції (glandulae endocrinae). Також цю групу залоз називають ендокринними залозами; вони анатомічно і топографічно різняться, мають різне походження. Залози внутрішньої секреції не мають вивідних проток і виділяють вироблені ними гормони (від грецького hormao – спонукати, збуджувати), чи інкрети, безпосередньо в кров, лімфу і міжклітинну рідину. Деякі гормони виробляють не окремі залози, а групи компактно розташованих клітин (підшлункові острівці, інтерстиціальні ендокриноцити яєчка – клітини Лейдіга), чи клітинами, що розміщені в різних органах і тканинах (клітини APUD-системи).
В організмі людини є дві складні системи керування функціями: нервова і гуморальна, котрі тісно пов’язані між собою і здійснюють єдину нейрогуморальну регуляцію. Центральна нервова система, включаючи кору півкуль великого мозку, регулює функції залоз внутрішньої секреції. Це здійснюється шляхом безпосередньої іннервації органів і тканин.
Гуморальна система (від латинського humor – волога; рідина) здійснює регулюючий вплив за допомогою гормонів, які потрапляють до органів і тканин через кровоносне і лімфатичне русла.
Гормони відрізняються від інших біологічно активних речовин такими ознаками:
– дія гормонів є дистантною, тобто органи, на які впливають гормони, віддалені від ендокринної залози;
– дія гормонів специфічна: одні гормони діють лише на визначені клітини-мішені, інші – на певні групи клітин;
– гормони мають високу біологічну активність і діють в дуже малих концентраціях;
– гормони діють тільки на живі клітини.
Гормони контролюють життєдіяльність організму, усі клітини, активність генів, формування клітинного фенотипу. Регулюючи активність ферментів, гормони впливають на обмін речовин (метаболізм).
Усю різноманітність дії гормонів можна об’єднати в три найважливіші функції: забезпечення росту і розвитку організму; забезпечення пристосування (адаптації) організму до умов середовища, що постійно змінюються; забезпечення гомеостазу – динамічної сталості внутрішнього середовища організму.
Залежно від ембріонального походження залози внутрішньої секреції поділяють на три групи – ентодермальні, мезодермальні та ектодермальні.
На сучасному етапі прийнято класифікувати ендокринні залози на залежні і незалежні від впливу передньої частки гіпофіза (аденогіпофіза). До першої групи, що залежать від гормонів аденогіпофіза, належать: щитоподібна залоза, кіркова речовина надниркових залоз, статеві залози (яєчка і яєчники).
До другої групи ендокринних залоз, на які безпосередньо не впливають гормони аденогіпофіза, належать: прищитоподібні залози, шишкоподібна залоза (шишкоподібне тіло), підшлункові острівці підшлункової залози, мозкова речовина надниркових залоз, параганглії.
Взаємодія між передньою часткою гіпофіза і незалежними від неї ендокринними залозами побудована за принципом прямих і непрямих зворотних зв’язків. Тропні гормони (від грецького tropos – напрямок) передньої частки гіпофіза (аденогіпофіза) активують діяльність гіпофіззалежних ендокринних залоз, а їхні гормони у свою чергу впливають на гіпофіз, пригнічуючи утворення і виділення відповідного гормону.Центром регуляції ендокринних функцій є гіпоталамус проміжного мозку, що координує нервові і гормональні механізми регуляції внутрішніх органів, об’єднує нервові й ендокринні регуляторні механізми в загальну нейроендокринну систему. Гіпоталамус утворює з гіпофізом єдиний функціональний комплекс, у якому перший виконує регулюючу, а другий – ефекторну функції. Отже, гіпоталамус і гіпофіз утворюють єдину гіпоталамо-гіпофізарну систему, що включає такі підсистеми:
– гіпоталамус–нейрогіпофіз (задня частка гіпофіза),
– гіпоталамус–аденогіпофіз (передня частка гіпофіза),
– гіпоталамо-гіпофізарну підсистему нейрорегуляторних пептидів.
Структура і функція гіпоталамуса та задньої частки гіпофіза докладно описані у розділі “Проміжний мозок”.
Гіпофіз
Гіпофіз (hypophysis), який ще називають мозковим придатком або (старовинна назва) пітуітарною залозою (glandula pituitaria), є найважливішою залозою внутрішньої секреції, що регулює діяльність ряду ендокринних залоз (щитоподібної, статевих, кіркової речовини надниркових залоз). Гіпофіз розміщений в гіпофізній ямці турецького сідла клиноподібної кістки. Відросток твердої оболони головного мозку – діафрагма сідла – відокремлює гіпофіз від порожнини черепа. Лійка, що проходить через діафрагму сідла, з’єднує гіпофіз із сірим горбом гіпоталамуса. Гіпофіз за формою нагадує квасолину, а його розміри у дорослої людини такі: поперечний – 10–17 мм, передньо-задній – 5–15 мм, вертикальний – 5–10 мм. Маса гіпофіза у чоловіків дорівнює 0,5–0,6 г, у жінок – 0,6–0,7 г.
Гіпофіз іззовні вкритий сполучнотканинною капсулою. Анатомічно єдиний гіпофіз складається з двох часток – передньої і задньої, які різняться за походженням.
Передня частка, або аденогіпофіз (lobus anterior; adenohypophysis) значно більша від задньої частки і займає 70–80 % усієї маси гіпофіза.
Аденогіпофіз складається з горбової частини (pars tuberalis), дальшої частини (pars distalis) і проміжної частини (pars intermedia). Менша задня частка, або нейрогіпофіз (lobus posterior; neurohypophysis) складається з лійки (infundibulum) і нервової частини (pars nervosa). Проміжна частина аденогіпофіза межує з задньою часткою.
Передня частка гіпофіза (аденогіпофіз) утворена тяжами епітеліальних клітин – ендокриноцитів, між ними розташовані сполучнотканинні прошарки, де проходять широкі синусоїдні капіляри. Між ендокриноцитами і базальною мембраною капілярів є перикапілярний простір. У передній частці є великі клітини, цитоплазматичні гранули яких інтенсивно зв’язують гістологічні барвники – це хромофільні аденоцити. Хромофобних аденоцитів більше, приблизно 60 %. Серед хромофільних аденоцитів розрізняють ацидофільні і базофільні ендокриноцити, які фарбуються відповідно кислими і основними гістологічними барвниками. В аденогіпофізі приблизно 40 % клітин є ацидофільними. Це круглі клітини середніх розмірів, у цитоплазмі яких містяться численні ацидофільні гранули. Великих базофільних клітин є приблизно 10 %. Інші клітини малі, слабо забарвлюються – це хромофобні аденоцити.
Ацидофільні ендокриноцити (альфа-клітини) виробляють два гормони – пролактин і соматотропін (ЛТГ і СТГ). Серед них є малі круглі клітини, в їхній цитоплазмі мало мітохондрій, а слабо виражений комплекс Гольджі синтезує пролактин. Соматотропні ендокриноцити, що синтезують соматотропін, круглої або овальної форми і середніх розмірів.
Усі базофільні ендокриноцити (бета-клітини) фарбуються основними гістологічними барвниками. Круглі гонадотропні ендокриноцити синтезують фолітропін і лютропін (ФСГ і ЛГ). Ядра клітин, що продукують фолітропін, круглі, а ядра лютропних клітин складчасті.
Великі неправильної форми тиротропні ендокриноцити, що виробляють тиротропін (ТТГ), мають велике видовжене ядро, розвинений комплекс Гольджі, багато широких мітохондрій і дрібних гранул неправильної форми діаметром 100–200 нм.
Кортикотропні ендокриноцити, що синтезують кортикотропін (АКТГ), неправильної форми, з широкими цитоплазматичними відростками, які закінчуються поблизу синусоїдних капілярів. Ядра цих клітин сегментовані.
Між ендокриноцитами є міжклітинні простори різної ширини. Аденоцити, що не синтезують гормонів, представлені дрібними клітинами з великим ядром і невеликим об’ємом цитоплазми.
Горбова частина аденогіпофіза пронизана малими артеріями, що кровопостачають передню частку, ворітними венами і синусоїдними капілярами. Між судинами розташовані тяжі епітеліальних клітин і псевдофолікули. Епітеліоцити представлені хромофобними, ацидофільними і базофільними аденоцитами, що мають типову для цього класу клітин будову.
Тонка проміжна частина аденогіпофіза утворена смужкою аденоцитів – меланотропних і ліпотропних, що виробляють секрет, який накопичується між клітинами, викликаючи ефект утворення псевдофолікулів. У сполучній тканині проміжної частини містяться поліморфні базальні клітини, амебоподібні відростки яких проникають по ходу судин у паренхіму задньої частки гіпофіза. З віком людини кількість таких відростків збільшується. У проміжній частині накопичуються меланотропін (МСГ), кортикотропін (АКТГ) і лютропін (ЛГ). Меланотропін впливає на пігментний обмін, а лютропін посилює обмін ліпідів. Проміжна частина гіпофіза пронизана пептидергічними нервовими волокнами (аксонами), що виходять з нейрогіпофіза. Ці аксони утворюють синапсоподібні контакти з епітеліоцитами. Холінергічні й амінергічні волокна гальмують секреторні процеси в аденоцитах.
Розвиток, вікові особливості та аномалії розвитку гіпофіза
Гіпофіз розвивається з двох різних зачатків: а) ектодермального випину ротової ямки зародка безпосередньо перед глотковою перетинкою, який має назву кишені Ратке, або гіпофізарної кишені; б) виросту проміжного мозку – гіпофізарної лійки. Починаючи з третього тижня ембріогенезу, кишеня Ратке поступово заглиблюється в дорсальному напрямку і наприкінці другого місяця зростається з лійкою, втрачаючи зв’язок з ротовою порожниною. В подальшому клітини передньої стінки кишені Ратке інтенсивно розмножуються і формують більшу частину передньої частки гіпофіза (аденогіпофіз). Невеликий виріст передньої стінки кишені Ратке утворює горбову частину аденогіпофіза, що росте вздовж лійки і поступово охоплює її. Задня стінка кишені Ратке дає початок проміжній частині аденогіпофіза. З виросту проміжного мозку (лійки) розвивається уся задня частка гіпофіза (нейрогіпофіз).
На дев’ятому тижні ембріогенезу починається диференціація базофільних ендокриноцитів, а на четвертому місяці пренатального онтогенезу – ацидофільних ендокриноцитів. У немовлят маса гіпофіза дорівнює приблизно 0,12 г, у 10-річних дітей його маса подвоюється, в 15-річних – потроюється, а у 20-річних людей вона максимальна. Після 60 років маса гіпофіза дещо зменшується. Упродовж перших 10–12 років життя в гіпофізі активно функціонують соматотропні і тиротропні ендокриноцити, у підлітків у гіпофізі значно зростає кількість гонадотропних ендокриноцитів.
Однією з аномалій розвитку гіпофіза є так званий горловий гіпофіз (скупчення клітин у вигляді стрічки довжиною 5–6 мм і товщиною 0.5–1 мм у підслизовій основі склепіння глотки), який утворюється з залишків кишені Ратке. З клітин горлового гіпофіза можуть розвинутися краніофарингіоми. Ці пухлини локалізуються в ділянці турецького сідла і спричинюють гідроцефалію та гіпофізарні дисфункції (наприклад, нецукровий діабет, порушення росту тощо).
Шишкоподібна залоза
Шишкоподібна залоза, або шишкоподібне тіло (glandula pinealis; corpus pineale) відома в медичній літературі як епіфіз (epiphysis), анатомічно належить до епіталамуса проміжного мозку (див. розділ “Проміжний мозок”).
Щитоподібна залоза
Щитоподібна залоза (glandula thyroidea) розташована в передній ділянці шиї попереду гортані і шийної частини трахеї, її маса у дорослої людини дорівнює 20–25 г. Щитоподібна залоза складається з двох часток – правої частки (lobus dexter) і лівої частки (lobus sinister), які поєднані між собою вузьким перешийком щитоподібної залози (isthmus glandulae thyroideae). Перешийок розташований на рівні дуги перснеподібного хряща гортані, а іноді на рівні 1–3 трахейних хрящів. У 30–35 % людей є ще пірамідна частка (lobus pyramidalis), яка відходить від перешийка залози вгору і може досягати тіла під’язикової кістки. Кожна частка має приблизно такі розміри: висоту – 5–6 см, ширину – 3–4 см, товщину – 2 см. Висота перешийка дорівнює 0,5–1,5 см, а товщина – 0,6–0,8 см. Щитоподібна залоза охоплює гортань спереду і з боків. Задньобокова поверхня кожної частки щитоподібної залози прилягає до гортанної частини глотки і переднього півкола загальної сонної артерії. Трапляються також додаткові щитоподібні залози (glandulae thyroideae accessoriae).
Щитоподібна залоза вкрита волокнистою капсулою (capsula fibrosa) і побудована зі строми (stroma) і паренхіми (parenchyma). Від волокнистої капсули, що складається з пучків колагенових і еластичних волокон, усередину залози відходять тонкі перегородки, поділяючи щитоподібну залозу на часточки (lobuli). У сполучнотканинних перегородках проходять судини і нерви. Паренхіма залози складається з мікроскопічних пухирців – фолікулів щитоподібної залози (folliculi glandulae thyroideae), що є структурно-функціональними одиницями залози .
У щитоподібній залозі є приблизно 30 млн фолікулів. Стінка фолікула утворена одним шаром кубічних клітин – тироцитів, розміщених на базальній мембрані. Діаметр фолікулів коливається від 0,05 до 0,5 мм. Трапляються розгалужені фолікули. Окрім того, окремі тироцити утворюють невеликі скупчення без порожнини всередині – міжфолікулярні острівці. Вважають, що тироцити острівців є джерелом утворення нових фолікулів. Форма тироцитів залежить від їх функціонального стану – при посиленні синтетичних процесів висота клітин збільшується. Фолікули обплетені густою сіткою кровоносних капілярів і сполучнотканинними волокнами.
У порожнині фолікулів накопичується колоїд – драглиста речовина, яку синтезують тироцити. Колоїд складається з білка тироглобуліну, в молекулі якого є гормон тироксин і поліпептидний ланцюг – глобулін.
Синтез і вивільнення гормонів із тироцитів є складним процесом. На рибосомах гранулярної ендоплазматичної сітки синтезується білок, який надходить в її цистерни, де до білка приєднуються бічні вуглеводні ланцюги. Білок переноситься транспортними пухирцями в комплекс Гольджі, де відбувається його подальше глікозування – утворення тироглобуліну. Від комплексу Гольджі відщеплюються просекреторні гранули, які перетворюються в секреторні, а згодом в апікальні пухирці. Апікальні пухирці з колоїдом виводяться з тироцитів у порожнину фолікула.
Йодування тироглобуліну відбувається в порожнині фолікула, в колоїді. З крові гемокапілярів у тироцити транспортується йод і надходить у колоїд, тут атомарний йод зв’язується з тирозином у складі тироглобуліну. Йодований тироглобулін накопичується в порожнині фолікула у формі колоїда. Крапельки колоїду фагоцитуються тироцитами. У цитоплазмі тироцитів ці крапельки з’єднуються з лізосомами, ферменти яких розщеплюють колоїд, унаслідок чого вивільняються гормони трийодтиро-нін (Т ) і тетрайодтиронін (Т ). Тетрайодтироніну є приблизно 90–95 % від усіх гормонів, які синтезують тироцити. Ці гормони виділяються через базальну поверхню тироцитів у перикапілярний простір, а з нього потрапляють у кров. Тироїдні гормони стимулюють окисні процеси в клітинах, регулюють водний, білковий, вуглеводний, жировий і мінеральний обміни; впливають на ріст, розвиток і диференціацію тканин.
Секрецію тироїдних гормонів стимулює гормон гіпофіза тиротропін (ТТГ), рівень якого регулює тироліберин гіпоталамуса. Тиротропін діє на тироцити шляхом активації аденілатциклази.
У стінці фолікулів, окрім тироцитів, наявний другий тип клітин – парафолікулярні ендокриноцити (кальцитоніноцити, або С-клітини). Це великі світлі клітини круглої або полігональної форми, які поодиноко залягають між основами тироцитів і базальною мембраною фолікулів (їхні верхівки не досягають просвіту фолікулів). У парафолікулярних клітинах є численні щільні мембранні аргірофільні секреторні гранули діаметром приблизно 0,15 мкм, які містять гормон кальцитонін. Цей гормон виділяється з клітин у перикапілярний простір, а згодом у кровоносні капіляри шляхом екзоцитозу. Кальцитонін регулює обмін кальцію і фосфору в організмі і є антагоністом паратгормону прищитоподібної залози. Цей гормон гальмує реабсорбцію кальцію з кісток і зменшує вміст кальцію в крові шляхом депонування його в кістковій тканині.
Кровопостачання та іннервація щитоподібної залози
Щитоподібна залоза інтенсивно кровопостачається, через неї проходить приблизно стільки ж крові, як через нирки. До верхніх кінців обох часток підходять права і ліва верхні щитоподібні артерії, які відходять від зовнішніх сонних артерій. До нижніх кінців часток підходять права і ліва нижні щитоподібні артерії з щито-шийних стовбурів підключичних артерій. Навколо щитоподібної залози формується непарне щитоподібне венозне сплетення, від якого венозна кров відтікає по правих і лівих верхній та середній щитоподібних венах у внутрішні яремні вени, а по нижніх щитоподібних венах у плечо-головні вени (чи в нижній відділ внутрішніх яремних вен).
Лімфатичні судини від щитоподібної залози впадають у глибокі передні шийні лімфатичні вузли: щитоподібні, передгортанні, передтрахейні і притрахейні.
Іннервують щитоподібну залозу нерви автономної частини периферійної нервової системи: симпатичні післявузлові волокна, що відходять від шийних симпатичних вузлів (переважно від середнього шийного вузла) і підходять до залози по ходу артерій; парасимпатичні та чутливі волокна проходять у складі верхнього гортанного і поворотного гортанного нервів, які є гілками блукаючого нерва (Х пара черепних нервів).
Розвиток, вікові особливості та аномалії розвитку щитоподібної залози
Щитоподібна залоза з’являється на четвертому тижні ембріогенезу як епітеліальний виріст нижньої частини глоткової кишки між компонентами зачатка язика (непарним горбиком і скобою) у точці, що відповідатиме сліпому отвору. Згодом щитоподібна залоза опускається спереду від глоткової кишки у вигляді роздвоєного дивертикула. Впродовж цієї міграції залоза залишається сполученою з язиком вузьким каналом – щитоязиковою протокою (ця протока пізніше зникає). Щитоподібна залоза досягає своєї кінцевої локалізації на сьомому тижні ембріогенезу. Вже тоді вона складається з маленького перешийка і двох часток. Щитоподібна залоза починає функціонувати приблизно наприкінці третього місяця, коли у ній з’являються перші фолікули, що містять колоїд.
У немовлят маса щитоподібної залози дорівнює 5–6 г, до першого року життя вона зменшується до 2–2,5 г. Потім маса залози поступово збільшується, досягаючи у дітей 12–14-річного віку 10–14 г, а до 25 років – 20–50 г. Після 65 років маса залози зменшується.
Порушення в процесі міграції щитоподібної залози в ембріогенезі може призвести до її неправильного положення (malpositio glandulae thyroideae), а саме: а) язикового положення; б) передтрахейного положення; в) загруднинного положення. Із залишків щитоязикової протоки може утворюватися щитоязикова кіста, яка іноді сполучається із зовнішнім середовищем каналом – щитоязиковою норицею (fistula thyrolingualis).
Прищитоподібні залози
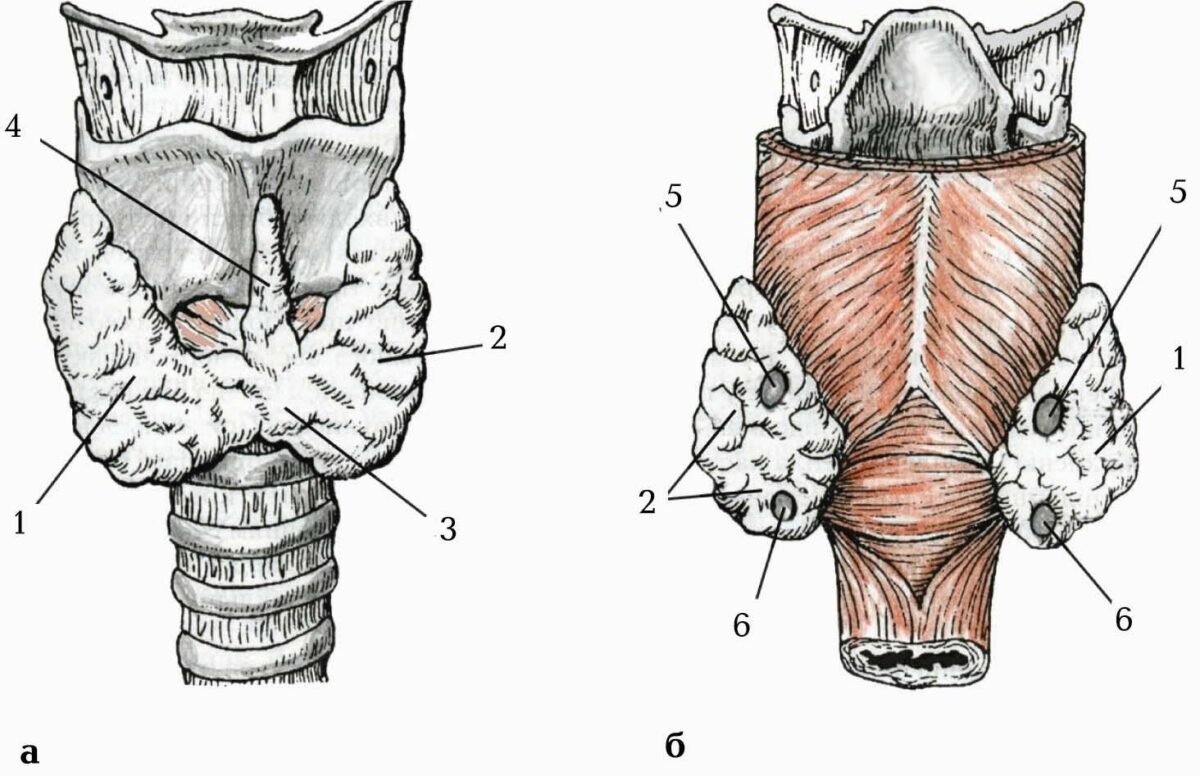
Дві пари невеликих прищитоподібних залоз (glandulae parathyroideae) розташовані на задній поверхні правої та лівої часток щитоподібної залози між її волокнистою капсулою і фасціальною пластинкою . Зазвичай з кожного боку є верхня прищитоподібна залоза (glandula parathyroidea superior) і нижня прищитоподібна залоза (glandula parathyroidea inferior). Верхня залоза розміщена на рівні межі між верхньою і нижньою третинами частки щитоподібної залози. Можлива наявність до восьми додаткових прищитоподібних залоз (glandulae parathyroideae accessoriae). Прищитоподібна залоза має вигляд видовженої горошини з такими розмірами: висота – 4–8 мм; ширина – 3–4 мм; товщина – 2–3 мм. Загальна маса всіх прищитоподібних залоз у дорослої людини дорівнює 0,20–0,35 г, а однієї залози – 0,05–0,09 г.
Як і всі ендокринні залози, кожна прищитоподібна залоза вкрита волокнистою капсулою, від якої всередину відходять тонкі сполучнотканинні пере-тинки, поділяючи паренхіму залози на ледь помітні часточки. У сполучній тканині капсули та перетинок є багато нервових волокон, кровоносних і лімфатичних судин.
Паренхіма прищитоподібної залози утворена епітеліальними клітинами – паратироцитами. Паратироцити, з’єднуючись між собою десмосомними контактами, утворюють перекладки (трабекули), що переплітаються. Перекладка (трабекула) є структурно-функціональною одиницею прищитоподібної залози. Іноді на гістологічних препаратах цієї залози у дітей видно, що паратироцити утворюють фолікулоподібні утвори, що нагадують фолікули щитоподібної залози. Розрізняють два види паратироцитів: головні паратироцити, цитоплазма яких забарвлюється базофільно (у залозі дітей до восьми років інших клітин немає), і ацидофільні паратироцити (утворюються в залозі дітей після 8–10-річного віку). Чітких закономірностей розташування клітин обох типів у паренхімі прищитоподібної залози не виявлено. Очевидно, обидва різновиди головних паратироцитів характеризують різні фази секреторної активності: синтезу гормона паратирину (паратгормону, паратиреоїдного гормону – ПТГ).
Паратгормон (паратирин) регулює концентрацію кальцію й опосередковано фосфору в крові, впливаючи таким чином на збудливість нервової і м’язової систем. Після видалення прищитоподібних залоз вміст кальцію в крові знижується, а фосфору підвищується. Паратгормон діє на кісткову тканину, активуючи остеобласти, обумовлюючи демінералізацію кісток і виділення іонів кальцію та фосфору в кров. Надлишок фосфору під впливом паратгормону виводиться нирками. Одночасно паратгормон посилює реабсорбцію іонів кальцію в канальцях нефронів, сприяючи зменшенню його виділення з сечею і підвищенню його вмісту в крові. Окрім того, паратгормон підвищує всмоктування іонів кальцію в кишці за умов надходження в організм необхідної кількості вітаміну D. Антагоністом паратгормону є гормон кальцитонін, що виробляється парафолікулярними клітинами (кальцитоніноцитами) щитоподібної залози. Він гальмує виведення кальцію з кісток, тому знижується його вміст у крові. Отже, клітини прищитоподібних залоз і парафолікулярні клітини щитоподібної залози реагують на зміну концентрації іонів кальцію в крові зміною інтенсивності синтезу і секреції відповідних гормонів, регулюючи таким чином вміст кальцію в крові.
Кровопостачання та іннервація прищитоподібних залоз
Прищитоподібні залози кровопостачають гілки верхніх і нижніх щитоподібних артерій, а також стравохідні і трахейні гілки (із зовнішніх сонних і підключичних артерій). Венозна кров від залоз відтікає у непарне щитоподібне венозне сплетення, що анастомозує з глотковими і гортанними венами, а потім у систему верхніх, середніх і нижніх щитоподібних вен.
Лімфатичні судини від прищитоподібних залоз впадають у глибокі передні шийні лімфатичні вузли: щитоподібні, передгортанні, передтрахейні і притрахейні.
Іннервують прищитоподібні залози гілки автономної частини периферійної нервової системи: симпатичні післявузлові волокна, що відходять від шийних симпатичних вузлів, підходять до залоз по ходу артерій; парасимпатичні та чутливі волокна проходять у складі верхнього гортанного і поворотного нервів, що є гілками блукаючого нерва (Х пара черепних нервів).
Розвиток, вікові особливості та аномалії розвитку прищитоподібних залоз
Прищитоподібні залози починають розвиток на сьомому тижні ембріогенезу з епітелію глоткової кишки (ентодерми) – з третьої і четвертої пар зябрових кишень. Епітеліальні зачатки залоз відокремлюються від стінок зябрових кишень, у процесі росту переміщуються каудально і займають постійне положення на задній поверхні правої та лівої часток щитоподібної залози.
У немовлят маса всіх прищитоподібних залоз не перевищує 10 мг, у однорічних дітей їхня маса досягає 20–30 мг, до 5 років – подвоюється, до 10 років – потроюється, а в 20-річному віці маса залоз досягає постійної величини, що не змінюється упродовж життя людини. Найбільша функціональна активність прищитоподібних залоз спостерігається у дітей віком 4–7 років.
З анатомії розвитку доволі часто зустрічається гетеротопія прищитоподібних залоз (нижня пара прищитоподібних залоз може бути розміщена у товщі щитоподібної залози, в передньому середостінні, в перикарді). Рідкісною аномалією є синдром Ді Джорджі (синдром ІІІ–ІV зябрових кишень), який включає гіпоплазію або аплазію прищитоподібних залоз та/або гіпоплазію або аплазію тимуса. Пацієнти з повним синдромом Ді Джорджі мають імунологічні проблеми, гіпокальцинацію та поганий прогноз у цілому. Синдром Ді Джорджі трапляється спорадично і може бути спричинений тератогенами, зокрема ретиноїдами.
Надниркові залози
Надниркова залоза (glandula suprarenalis) парна (права і ліва), за формою нагадує сплощену спереду назад піраміду з заокругленою верхівкою . Залози розташовані в заочеревинному просторі на рівні ХІ–ХІІ грудних хребців у товщі жирової капсули кожної нирки, прилягаючи до верхньоприсередньої ділянки верхнього кінця нирки, її оточує ниркова фасція. Права надниркова залоза розміщена дещо нижче від лівої залози. Маса надниркової залози в дорослої людини дорівнює приблизно 12–15 г, а її розміри такі: довжина – 40–60 мм; товщина (передньо-задній розмір) – 5–10 мм; висота – 20–30 мм. Надниркова залоза має три поверхні: передню поверхню (facies anterior), задню поверхню (facies posterior) і нижню ввігнуту – ниркову поверхню (facies renalis), а також два краї – верхній край (margo superior) і присередній край (margo medialis). Поверхні залоз дещо горбисті.
У людини можлива наявність додаткових надниркових залоз (glandulae suprarenales accessoriae).
Права надниркова залоза спереду має вигляд трикутника з заокругленими кутами. Його задня поверхня прилягає до поперекової частини діафрагми, нижня частина передньої поверхні залози, що вкрита очеревиною, прилягає до нутрощевої поверхні печінки і дванадцятипалої кишки. Ниркова поверхня надниркової залози охоплює верхній кінець правої нирки, а присередній край залози прилягає до нижньої порожнистої вени.
Ліва надниркова залоза має півмісяцеву форму, її ниркова поверхня заходить на верхню частину присереднього краю лівої нирки, присередній край залози прилягає до черевної аорти. Передня поверхня лівої надниркової залози прилягає до хвоста підшлункової залози і кардіальної частини шлунка, а задня поверхня залози стикається з поперековою частиною діафрагми.
На передній поверхні надниркової залози є глибока борозна – ворота (hilum). У правій залозі ворота розташовані ближче до її верхівки, а в лівій залозі – ближче до основи. Через ворота залози виходить центральна вена (v. centralis).
Надниркова залоза вкрита тонкою волокнистою капсулою, в якій містяться гладкі міоцити. Від капсули вглиб залози відходять тонкі сполучнотканинні перетинки, розділяючи її кіркову речовину на численні епітеліальні тяжі, обплетені густою сіткою гемокапілярів.
Паренхіма надниркової залози складається з двох шарів, що походять з різних зародкових листків. Зовнішній шар називається кірковою речовиною (cоrtex), а внутрішній шар – мозковою речовиною (medulla).
Кіркова речовина розвивається з мезодерми на п’ятому тижні ембріогенезу, а саме – з целомічного епітелію спланхнотома між коренем дорсальної брижі первинної кишки і сечово-статевою складкою. Тканина, що розвивається з мезодерми, розміщується між двома первинними нирками, тому називається інтерреналовою тканиною (інтерреналовим тілом). Із цього ж джерела утворюються зачатки статевих залоз і додаткових надниркових залоз.
Мозкова речовина надниркових залоз, як і нервова система, має ектодермальне походження. Вона починає формуватися на шостому-сьомому тижнях ембріогенезу з зачатків вузлів симпатичного стовбура. Нервові клітини – симпатобласти – відокремлюються від симпатичних вузлів і перетворюються на хромафінобласти, які в свою чергу диференціюються на хромафіноцити (забарвлюються солями хрому). Ці клітини переміщуються в інтерреналове тіло, утворюючи мозкову речовину надниркової залози, яку охоплює кіркова речовина. Так формується анатомічно єдина залоза. Із симпатобластів формуються інші хромафінні тільця – параганглії.
У кірковій речовині виділяють три зони : клубочкову (зовнішню), пучкову (середню) і сітчасту (внутрішню, яка межує з мозковою речовиною). Співвідношення товщини цих зон відповідно дорівнює 1: 9: 3. Вони різняться між собою як структурно, так і функціонально.
Клубочкова зона (zona glomerulosa) складається з малих призматичних клітин, які утворюють скупчення у вигляді клубочків. Клубочки оточені сіткою звивистих гемокапілярів. Ендокриноцити клубочкової зони продукують мінералокортикостероїдні гормони, переважно альдостерон.
Пучкова зона (zona fasciсulata), яка є найтовстішою, побудована з великих світлих полігональних клітин, які формують довгі тяжі – пучки. Ці пучки орієнтовані перпендикулярно до поверхні органа. Між пучками проходять прямі гемокапіляри, анастомозуючи між собою. Ендокриноцити пучкової зони виробляють глюкокортикостероїдні гормони – кортизол (гідрокортизон) і кортикостерон. Ці клітини синтезують також незначну кількість андрогенів – дегідроепіандростерон (дегідроізоандростерон) і андростендіон.
У сітчастій зоні (zona reticularis) клітини полігональної або круглої форми (дещо менші від ендокриноцитів пучкової зони) утворюють розгалужені тяжі, що нагадують сітку. Між групами клітин проходять гемокапіляри. Ендокриноцити сітчастої зони синтезують стероїдні гормони – андрогени, естрогени і прогестерон, а також незначну кількість глюкокортикоїдів.
Отже, в кірковій речовині надниркової залози її структурно-функціональні зони гістологічно чітко розмежовані між собою. Ендокриноцити цих зон синтезують різні за функцією гормони: клубочкова зона – мінералокортикоїди (альдостерон); пучкова зона – глюкокортикостероїди (кортизол, кортикостерон); сітчаста зона – стероїдні гормони (андрогени, естрогени, прогестерон).
Мінералокортикоїди беруть участь в регуляції електролітного обміну й опосередковано – в регуляції водного обміну. Альдостерон посилює реабсорбцію іонів натрію в нирках, слинних залозах і кишковошлунковому тракті, тобто затримує іони натрію в організмі. Альдостерон змінює проникність клітинних мембран для іонів натрію і калію, посилює запальні процеси й утворення колагену. Секрецію альдостерону ендокриноцитами регулює концентрація іонів натрію і калію в плазмі крові й ангіотензин
ІІ. Збільшення концентрації іонів калію і зменшення іонів натрію, а також зменшення об’єму плазми крові приводить до посилення секреції альдостерону. Подібний ефект дає ангіотензин ІІ, що утворюється з ангіотензину І плазми крові під впливом гормону реніну, який виробляється юкстагломерулярними клітинами нирки.
Глюкокортикоїди сильно і різнобічно впливають на обмінні процеси в організмі людини, посилюючи катаболічні процеси при білковому обміні і стимулюючи глюконеогенез, що призводить до підвищення рівня глюкози в крові і глікогену в печінці, скелетних м’язах і міокарді. Глюкокортикоїди регулюють ліполіз, нормалізують виділення води з організму, підвищують клубочкову фільтрацію і зменшують реабсорбцію води в дистальних звивистих канальцях нирки. У лікарській практиці широко використовується протизапальна дія глюкокортикоїдів. Введення гідрокортизону викликає інволюцію імунної системи, виражену імуносупресію, пов’язану з руйнуванням лімфоїдної тканини.
Глюкокортикоїди вибірково гальмують утворення основної речовини сполучної тканини і проліферацію фібробластів, зменшують кількість тканинних базофілів, блокують дію ферменту гіалуронідази і тим самим зменшують проникність капілярів. Стрес-фактори викликають збільшення секреції глюкокортикоїдів навіть до повного “виснаження” ендокриноцитів кіркової речовини надниркових залоз. Секреція глюкокортикоїдів ендокриноцитами пучкової і частково сітчастої зон регулюється кортикотропіном (адренокортикотропним гормоном – АКТГ), який секретується аденогіпофізом.
Мозкова речовина надниркової залози відмежована від кіркової речовини тонким несуцільним сполучнотканинним прошарком. Її паренхіма утворена скупченнями великих круглих або полігональних клітин, розділених синусоїдними гемокапілярами і посткапілярними венулами . Ці клітини добре забарвлюються солями хрому в коричневий колір, тому вони називаються хромафінними клітинами. До клітин підходять передвузлові симпатичні нервові волокна, утворюючи з ними синапси. За характером синтезованих речовин у мозковій речовині надниркової залози розрізняють два види клітин: епінефроцити, що виробляють адреналін, і норепінефроцити, що синтезують норадреналін. Адреналін і норадреналін об’єднані в групу біологічно активних речовин – катехоламінів.
Клітини мозкової речовини надниркових залоз продукують невелику кількість гормонів, хоча при впливі на організм сильних подразників їх секреція різко зростає.
У дорослої людини мозкова речовина надниркових залоз виробляє приблизно 70–90 % адреналіну і 10–30 % норадреналіну. Молекули катехоламінів зв’язують специфічні рецептори цитолеми клітинмішеней. Норадреналін викликає звуження судин (крім судин головного мозку), а адреналін звужує певні судини (судини шкіри) і розширює інші (вінцеві судини серця, судини скелетних м’язів). Адреналін підвищує систолічний артеріальний тиск, дещо знижуючи діастолічний тиск. Норадреналін підвищує систолічний і діастолічний тиск. Обидва гормони розслаблюють гладкі м’язи бронхів, сприяючи інтенсивнішому диханню.
Адреналін підвищує рівень цукру в крові, викликаючи розщеплення глікогену. Ідентичний ефект норадреналіну слабший. Адреналін і норадреналін посилюють розщеплення жирів, активують енергетичний обмін. Секреція катехоламінів у звичайних фізіологічних умовах невелика і дорівнює 8–10 нг/ (кг-хв.), але різко зростає в умовах стресу.
Кровопостачання та іннервація надниркових залоз
Кожну надниркову залозу кровопостачає 25–30 артерій, але найкрупнішими є три артерії: верхня надниркова артерія (від нижньої діафрагмової артерії); середня надниркова артерія (від черевної частини аорти); нижня надниркова артерія (від ниркової артерії). Гілки одних артерій кровопостачають тільки кіркову речовину, інші пронизують кіркову речовину і галузяться в мозковій речовині. Важливим є те, що клітини мозкової речовини контактують як з гемокапілярами, так і з посткапілярними венулами, в які кров поступає з капілярів. Синусоїдні гемокапіляри формують притоки центральної вени, яка виходить із залози на її передній поверхні через ворота. Після виходу з залози центральна вена називається наднирковою веною. Ліва надниркова вена впадає в ліву ниркову вену, а права надниркова вена – безпосередньо в нижню порожнисту вену. Численні дрібні вени від надниркових залоз впадають в притоки ворітної печінкової вени.
Лімфатичні судини, відходячи від надниркових залоз, впадають у поперекові лімфатичні вузли, які розміщені навколо аорти і нижньої порожнистої вени. В органі лімфатичні судини містяться в капсулі і поблизу крупних вен, де є прошарки сполучної тканини.
Надниркові залози іннервує автономна частина периферійної нервової системи – симпатичні і парасимпатичні волокна, які утворюють надниркове нервове сплетення. Парасимпатичні і чутливі волокна проходять у складі гілок блукаючого нерва (Х пара черепних нервів). Унікальною є іннервація мозкової речовини надниркових залоз, яку здійснюють тільки передвузлові симпатичні волокна. Від бічної частини черевного симпатичного вузла (відповідно правого і лівого) до надниркової залози відходить приблизно 20 гілочок, які беруть участь в утворенні надниркового нервового сплетення. У складі цих гілок проходять передвузлові симпатичні волокна, безпосередньо іннервуючи ендокриноцити мозкової речовини залози.
Вікові особливості надниркових залоз
У немовлят маса одної надниркової залози дорівнює приблизно 8–9 г і значно більша за масу залози у дитини першого року життя. Упродовж перших трьох місяців після народження маса надниркової залози різко зменшується до 3–4 г, бо в цей період стоншується і зазнає перебудови кіркова речовина. Згодом маса залози поступово збільшується. У 5-річних дітей маса залози знову дорівнює 8–9 г, як і в немовлят. Формування кіркової речовини надниркової залози завершується в період другого дитинства (8–12 років). До 20 років маса кожної залози збільшується в 1,5 раза (у порівнянні з немовлятами) і досягає максимуму – 12–13 г. У наступні вікові періоди розміри і маса надниркових залоз майже незмінні. У жінок маса надниркових залоз дещо більша, ніж у чоловіків. Під час вагітності маса кожної залози збільшується приблизно на 2 г. Тільки після 70-річного віку у людей відбувається незначне зменшення розмірів і маси надниркових залоз.
Параганглії
Крім мозкової речовини надниркових залоз, хромафіноцити містяться в парагангліях (paraganglia), що утворюються на сьомому тижні ембріогенезу з зачатків симпатичної частини нервової системи і тісно пов’язані з симпатичними вузлами автономного відділу периферійної нервової системи. Більшість парагангліїв розташовані позаочеревинно поблизу симпатичного стовбура у вигляді невеликих відокремлених структур . Хромафіноцити парагангліїв, як і мозкової речовини надниркових залоз, синтезують катехоламіни – адреналін і норадреналін.
Найбільшими парагангліями є парний сонний клубочок (glomus caroticum) і приаортові тіла, або аортові клубочки (corpora paraaortica; glomera aortica). Численні дрібні параганглії розташовані позаочеревинно в ділянках від надниркових до статевих залоз, а також по ходу симпатичних нервів і в товщі симпатичних вузлів симпатичні параганглії (paraganglia sympathica). Вони розташовані поблизу пухирчастих залоз у чоловіків, а у жінок – у матково-піхвовому нервовому сплетенні. Кількість і розміри парагангліїв дуже мінливі. У немовлят є понад 40 парагангліїв, а також численні скупчення хромафіноцитів і окремі хромафіноцити у симпатичних вузлах та по ходу симпатичних нервів.
Параганглії оточені тонкою сполучнотканинною капсулою, а її волокна проникають вглиб паренхіми. Паренхіма парагангліїв складається з хромафіноцитів, які утворюють тяжі, що переплітаються між собою. У просвітах між клітинними тяжами проходять численні гемокапіляри, хромафіноцити оточені густою сіткою симпатичних передвузлових волокон. Іннервація парагангліїв подібна до іннервації мозкової речовини надниркових залоз. Кровопостачають параганглії гілочки, що відходять від сусідніх найменших артерій. Інволюція парагангліїв починається у півторарічних дітей і закінчується в юнацькому віці.
Сонний клубочок (glomus caroticum) – парний веретеноподібний утвір сіро-рожевого кольору, довжиною 5–8 мм, шириною 1,5–5 мм і товщиною 1–1,5 мм. У дітей довжина сонного клубочка не перевищує 2 мм. Сонний клубочок розміщений в основі роздвоєння сонної артерії, заходячи на задньоприсередню поверхню загальної сонної артерії.
По ходу аорти розміщується більшість парагангліїв – надсерцеві параганглії, приаортові тіла і куприковий клубочок.
Надсерцеві параганглії (paraganglia supracardiales) представлені верхнім і нижнім надсерцевими парагангліями (paraganglia supracardiales superior et inferior). Нижній параганглій непостійний, з віком він редукується. Верхній надсерцевий параганглій більший, розташований між легеневим стовбуром і висхідною частиною аорти на межі з її дугою. Нижній надсерцевий параганглій залягає в ділянці виходу з аорти лівої вінцевої артерії.
Приаортові тіла, або аортові клубочки (corpora paraaortica; glomera aortica) складаються з правої та лівої частин. Кожна частина має вигляд тяжа, що залягає на бічній поверхні черевної аорти. Довжина правої частини становить 8–20 мм, лівої частини – 8–15 мм, товщина дорівнює 2–3 мм. У дітей першого року життя аортові клубочки найбільші: у немовлят довжина приаортових тіл дорівнює 8–15 мм, а товщина 2–3 мм, у дітей грудного віку довжина цих утворів досягає 30 мм. Обидва тяжі інколи з’єднані між собою смужкою хромафінних клітин, розміщених попереду аорти. Приаортові тіла зв’язані з гілками нижніх грудних і поперекових вузлів симпатичного стовбура.
Куприковий клубочок (glomus coccygeum) непарний, розташований на передній поверхні верхівки куприка і має довжину приблизно 2,5 мм.
Підшлункові острівці
Підшлункова залоза складається з ендокринної та екзокринної частин. Екзокринна частина описана в розділі “Травна система”. Ендокринна частина представлена підшлунковими острівцями (insulae pancreaticae) – острівцями Лангерганса, що сформовані зі скупчень ендокринних клітин – інсулоцитів . Інсуліноцити, як і весь острівець, оточені густою сіткою гемокапілярів фенестрованого типу. Між інсулоцитами і гемокапілярами є перикапілярні простори, тому гормони, які виробляють інсулоцити, виділяються у цей простір, а потім через стінку капілярів потрапляють у кров. У підшлунковій залозі є приблизно 1–2 млн підшлункових острівців, діаметр кожного з яких дорівнює 100–300 мкм. Найбільша кількість підшлункових острівців у хвості залози.
Інсулоцити – це невеликі клітини з цитоплазмою, яка слабо фарбується барвниками, тому на гістологічних препаратах підшлункові острівці виглядають світлішими на тлі темніших екзокринних панкреацитів. У цитоплазмі інсулоцитів добре розвинений комплекс Гольджі і помірно гранулярна ендоплазматична сітка, є багато великих мітохондрій. Наявність численних мембранних гранул у цитоплазмі інсулоцитів є їх характерною ознакою. Ці гранули суттєво відрізняються від гранул зимогену ацинозних клітин. За структурними і хімічними властивостями гранул інсулоцити поділяють на чотири основні групи: В-клітини (базофільні), А-клітини (ацидофільні), D-клітини (дендритні), F-клітини.
У підшлункових острівцях найбільше В-інсулоцитів – 70–75 %, які розміщені переважно в центрі острівців. Вони містять численні дрібні базофільні гранули діаметром приблизно 275 нм, що специфічно забарвлюються альдегід-фуксином у фіолетовий колір. У гранулах міститься гормон – інсулін. Між мембраною гранули та її вмістом є світлий прошарок.
Вміст кожної гранули складається з одного-декількох прямокутних чи полігональних кристалоїдів, що занурені в матрикс. Ядра неправильної форми, розташовані в центрі В-клітин. На рибосомах гранулярної ендоплазматичної сітки синтезується проінсулін, що піддається процесингу в комплексі Гольджі. Від нього відщеплюються секреторні гранули, що містять інсулін. Гормон інсулін збільшує проникність клітинних мембран для глюкози, яка присутня у плазмі крові, активує утворення включень глікогену в клітинах. Цими властивостями пояснюється гіпоглікемічна дія інсуліну. При зменшенні концентрації інсуліну в крові клітини не “споживають” глюкозу, тому її рівень в крові різко підвищується і глюкоза потрапляє в сечу. В-інсуліноцити синтезують також гормон амілін, який бере участь у регуляції вуглеводного обміну.
А-інсулоцити складають 20–25 % від усіх клітин острівця, розташовані здебільшого групами на периферії підшлункових острівців. Ядра – неправильної форми розміщені в центрі клітин, гетерохроматину небагато. У цитоплазмі А-інсулоцитів розміщені численні круглі щільні оксифільні гранули діаметром 230–300 нм, що оточені мембранами. Щільний вміст гранул відокремлений від їх мембран вузьким світлим обідком. Гранули забарвлюються кислим фуксином у червоний колір. У цитоплазмі є невелика кількість малих мітохондрій, слабо розвинений комплекс Гольджі і помірна кількість коротких цистерн гранулярної ендоплазматичної сітки. В гранулах А-інсулоцитів міститься гормон глюкагон, який є антагоністом інсуліну. Глюкагон активує глікогеноліз і ліполіз, при цьому глікоген розщеплюється до глюкози, а її рівень в крові підвищується.
D-інсулоцити мають великі розміри і зірчасту форму, їх приблизно 5–10 %. У цитоплазмі містяться великі щільні гранули без світлого обідка діаметром 320–350 нм. У цих клітинах органел значно менше, ніж у Аі В-клітинах. Д-клітини синтезують гормон соматостатин, який гальмує синтез інсуліну і глюкагону відповідно Ві А-інсулоцитами, синтез ферментів ациноцитами підшлункової залози і соматотропіну (гормону росту) аденогіпофізом.
F-інсулоцитів дуже мало – всього 1–5 %, вони містять дрібні гранули діаметром приблизно 140 нм і розташовані на периферії підшлункових острівців. F-клітини виробляють поліпептид – стимулятор виділення шлункового соку і підшлункового соку екзокринною частиною підшлункової залози.
Навколо підшлункових острівців наявні також невеликі групи секреторних клітин, що називаються проміжними клітинами. Вони містять найдрібніші щільні аргірофільні секреторні гранули діаметром приблизно 100 нм зі світлим обідком. Вважають, що ці клітини синтезують трипсиноподібні ферменти, які вивільняють інсулін з проінсуліну, а також виділяють вазоактивний поліпептид.
Кровопостачання та іннервація підшлункових острівців
Кожен підшлунковий острівець кровопостачають 1–3 артеріоли, що відходять від дрібних артеріальних гілок (див. кровопостачання підшлункової залози). Від артеріол відходять численні капіляри фенестрованого типу, які проходять між інсулоцитами. Інсулоцити виділяють гормон спочатку в перикапілярний простір, а згодом через стінку капілярів потрапляють у кров. Капіляри збираються у венули, які формують вени. Венозна кров по підшлунково-залозових венах прямує у селезінкову вену, верхню брижову вену та інші притоки ворітної печінкової вени. Капілярні сітки екзокринних і ендокринних компонентів не сполучаються між собою. У підшлункових острівців лімфатичні капіляри відсутні.
Підшлункові острівці іннервують симпатичні і парасимпатичні волокна автономної частини периферійної нервової системи. Ці волокна утворюють навколо острівців сплетення, гілочки яких проникають в острівці. По парасимпатичних волокнах переважно правого блукаючого нерва (Х пара черепних нервів), в інтрамуральних вузлах якого розташовані холінергічні нейрони, передається команда на посилення секреції інсуліну В-клітинами підшлункових острівців. По чутливих волокнах блукаючого нерва передається чутлива інформація від острівців. По післявузлових симпатичних волокнах, що відходять від черевних симпатичних вузлів, передається команда на посилення секреції глюкагону А-клітинами
Ендокринна частина статевих залоз
Статеві залози – яєчка у чоловіків і яєчники у жінок – виробляють статеві гормони, що потрапляють у кров. Як сказано вище (див. розділ підручника “Cтатеві системи”), цю функцію в яєчках здійснюють інтерстиційні ендокриноцити (клітини Лейдіга). Це великі клітини, скупчення яких розташовані між звивистими сім’яними трубочками біля кровоносних капілярів. Ендокриноцити яєчка беруть активну участь в утворенні чоловічого статевого гормону – тестостерону (андрогену), що діє на різні андрогенчутливі клітини чоловічого організму, стимулюючи їх ріст та функціональну активність. До таких клітин-мішеней належать клітини передміхурової залози, пухирчастих (сім’яних) залоз, залоз передньої шкірочки статевого члена, нирок і шкіри. Під впливом андрогенів відбувається ембріональна диференціація і розвиток чоловічих статевих органів, статеве дозрівання і розвиток вторинних статевих ознак, опорно-рухового апарату. Разом з іншими гормонами андрогени регулюють ріст кісток у довжину і товщину, а після досягнення організмом статевої зрілості їх дія слабшає. Андрогени стимулюють синтез білків і прискорюють ріст тканин. Дуже важливим є їх вплив на сперматогенез: низька концентрація андрогену активує цей процес, висока – гальмує. Андрогени зумовлюють статеву поведінку чоловіків, їхнє лібідо (статеве бажання; потяг) і потенцію. Варто пам’ятати, що в яєчках синтезується невелика кількість естрогенів – жіночих статевих гормонів.
Жіночі статеві гормони виробляють яєчники. Клітини фолікулярного епітелію (зернистого шару фолікулів) синтезують естроген (фолікулін, або естрадіол), клітини жовтого тіла – лютеоцити – продукують гестаген (прогестерон). Крім того, в яєчниках утворюється невелика кількість чоловічих статевих гормонів – андрогенів. Естрогени, як і андрогени в чоловіків, впливають на ембріональне диференціювання і розвиток жіночих статевих органів, вторинних статевих ознак, ріст і розвиток опорно-рухового апарату; забезпечують розвиток тіла за жіночим типом, впливають на статеву поведінку жінок. Жіночі статеві гормони регулюють описані вище зміни ендометрію під час менструального циклу, стимулюють проліферацію ендометрію. Прогестерон впливає на слизову оболонку матки (ендометрій), готуючи її до імплантації зародка і розвитку плода, грудей, а також затримує ріст нових фолікулів в яєчниках. Окрім того, статеві гормони мають і екстрагенітальну дію. Так, прогестерон викликає підвищення ранкової температури тіла в стані спокою (базальної температури), має катаболічну дію.
Дифузна нейроендокринна система (АPUD-СИСТЕМА)
В організмі людини наявні гормонопродукуючі клітини – ендокриноцити, що синтезують гормональні нейроаміни і олігопептиди. Цей тип клітин розвивається з нейробластів нервового гребінця, а також з ектодерми і ентодерми, вони розсіяні в органах і системах організму у вигляді окремих ендокриноцитів (апудоцитів) чи клітинних скупчень. Всі ці клітини об’єднані в APUD-систему (Amine Precursors Uptake and Decarboxylation – поглинання і декарбоксилування попередників амінів).
До APUD-системи належать: парафолікулярні клітини щитоподібної залози, клітини мозкової речовини надниркових залоз; нейросекреторні клітини гіпоталамуса; пінеалоцити шишкоподібної залози; головні паратироцити прищитоподібних залоз; ендокриноцити аденогіпофіза, плаценти, підшлункової залози і шлунково-кишкового тракту.
Окремо виділено групу дисоційованих клітин ненейрального походження, що не нагромаджують і не декарбоксилують попередники біологічно активних амінів, а саме: ендокриноцити яєчок, фолікулярні клітини і лютеоцити яєчників. Ці клітини продукують стероїдні гормони – тестостерон, естрогени, прогестерон. Їхню функцію регулює відповідний тропний гормон гіпофіза.
Отже, гормони APUD-системи забезпечують як місцеву (паракринну), так і дистантну регуляцію діяльності органів і систем організму. Функція клітин APUD-системи не залежить від дії гормонів гіпофіза, але регулюється нервовими імпульсами, які передаються по симпатичних і парасимпатичних волокнах автономної частини периферійної нервової системи. APUD-система доповнює і об’єднує між собою нервову й ендокринну системи, забезпечуючи регуляцію гомеостазу.