Генетика людини: методичка
2. ОСНОВИ ЗАГАЛЬНОЇ ГЕНЕТИКИ
2.1. Генетика якісних ознак
Усі ознаки живих організмів, які вивчає генетика, умовно можна поділити на якісні та кількісні. Якісними ознаками є такі ознаки, які можуть бути або наявними, або відсутніми, наприклад, забарвлення квітки рослини: наявне – червоні пелюстки або відсутнє – білі пелюстки. Якісні ознаки називають також альтернативними. Кількісні ознаки характеризуються непереривною мінливістю, наприклад, зріст людини. Такі ознаки визначають вимірюванням.
Сучасна генетика була започаткована дослідженнями якісних ознак, бо закономірності їх успадкування досить прості та більш доступні для генетичного аналізу. Тому ознайомлення з основами генетики ми і розпочнемо з викладу понять та закономірностей успадкування цих ознак.
2.1.1. Закони Менделя
Перші генетичні дослідження. Довгий час матеріальний носій спадковості сприймався як однорідна за складом речовина. Вважалося, що спадкова речовина батьків змішується у дитини подібно двом взаєморозчинним рідинам. Відповідно до цього гібридне потомство, одержане в результаті з’єднання спадкового матеріалу організмів, що розрізняються за рядом ознак, повинно мати проміжні значення цих ознак. І дійсно, такі явища часто спостерігаються. У людини та тварин речовину спадковості пов’язували з кров’ю. Звідси відомі вирази – «напівкровний», «чистокровний», які збереглися до наших днів.
Проте у другій половині XIX століття деякі дослідники спостерігали у гібридів таку мінливість ознак, яку не можна було пояснити з позиції вищезгаданого уявлення. Одним з цих дослідників був Грегор Мендель. Він першим показав, що спадкові чинники не змішуються, а передаються з покоління в покоління у вигляді незмінних окремих одиниць. Ці спадкові одиниці передаються через чоловічі та жіночі статеві клітини – гамети. У кожної особини спадкові одиниці перебувають у вигляді пар, тоді як у гаметах наявна лише одна одиниця з кожної пари. Ці одиниці спадковості Г. Мендель назвав елементами.
У 1900 році, коли закони Менделя були відкриті вдруге і одержали визнання, одиниці спадковості були названі факторами. Через декілька років датський біолог В. Йогансен дав їм назву «гени», а в 1912 році американський генетик Т.Г. Морган показав, що гени розташовані в хромосомах.
Успіх Г. Менделя багато в чому обумовлений вдалим вибором експериментального об’єкту – гороху. Ця рослина має ряд сприятливих властивостей для проведення експериментів по схрещуванню. По-перше, сорти гороху чітко розрізняються за рядом простих, якісних ознак. По-друге, горох – самозапильна рослина, завдяки чому підтримується чистота сорту, тобто збереження ознаки від покоління до покоління. По-третє, можна схрещувати рослини шляхом штучного запилення і одержувати потрібні гібриди. Гібриди гороху здатні давати потомство, що при гібридизації інших рослин зустрічається не завжди.
Г. Менделю вдалося підібрати такі пари альтернативних ознак, які, як це було встановлено пізніше, мають простий тип успадкування: форма насіння (гладенька або зморшкувата), забарвлення насіння (жовте або зелене), забарвлення квіток (біле або кольорове) та деякі інші.
Подібні досліди з гібридизації рослин неодноразово проводилися і до Г. Менделя, але ніхто не зміг одержати таких всеохоплюючих даних, а головне, побачити в них закономірності спадковості. Варто особливо зупинитися на тих моментах, які забезпечили Г. Менделю успіх, оскільки його дослідження можна вважати зразком проведення будь-якого наукового експерименту. Перш ніж почати основні експерименти, Г. Мендель провів попереднє вивчення експериментального матеріалу, після чого ретельно спланував усі експерименти. Основним принципом його дослідження була поетапність – спочатку вся увага була спрямована на одну ознаку, що спрощувало аналіз, а потім аналізувалася наступна ознака. Одержувані дані ретельно реєструвалися. Г. Мендель провів безліч експериментів і одержав достатню кількість даних для забезпечення статистичної достовірності результатів.
Моногібридне схрещування. Перший та другий закони Менделя. Вивчаючи результати схрещування рослин з альтернативними ознаками (наприклад, насіння: гладеньке – зморшкувате, квітки: білі – забарвлені), Г. Мендель виявив, що гібриди першого покоління (F1), одержані за допомогою штучного запилення, не є проміжними між двома батьківськими формами, а в більшості випадків схожі на одну з них. Наприклад, при схрещуванні рослин із забарвленими та білими квітками все потомство першого покоління мало забарвлені квітки (мал. 1). Та батьківська ознака, якою володіли рослини першого покоління, Г. Мендель назвав домінантною. У наведеному прикладі домінантною ознакою буде наявність забарвлення у квіток.
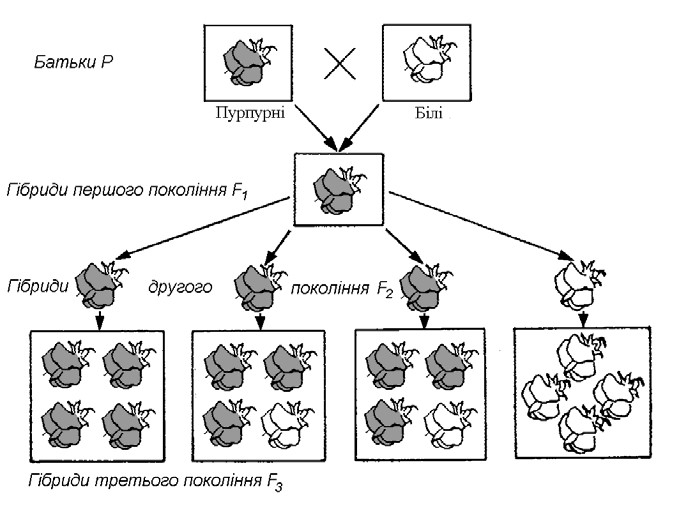
Мал. 1. Успадкування пурпурного та білого забарвлення квіток у гороху
Від експериментальних гібридів (F1) шляхом самозапилення Г. Мендель одержав потомство другого покоління (F2) і виявив, що воно не одноманітне: частина з них несе ознаку тієї батьківської рослини, яка не проявилася у гібридів першого покоління. Отже, ознака, відсутня в поколінні F1, знову з’явилася в поколінні F2. Г. Мендель зробив висновок, що ця ознака була присутня в поколінні F1 у прихованому вигляді. Цю ознаку він назвав рецесивною. У наведеному прикладі рецесивною ознакою буде біле забарвлення квіток.
Г. Мендель здійснив цілу серію подібних дослідів із різними парами альтернативних ознак і кожного разу ретельно підраховував співвідношення рослин з домінантними та рецесивними ознаками. У всіх випадках аналіз показав, що відношення кількості рослин з домінантною ознакою до кількості рослин з рецесивною у поколінні F2 складало приблизно 3 : 1.
У третьому поколінні (FЗ), одержаному також шляхом самозапилення рослин покоління F2, виявилося, що ті рослини з другого покоління, які несли рецесивну ознаку, дали потомство, всі рослини якого мали цю ознаку. Частина рослин F2 з домінантною ознакою дала потомство з цією ж ознакою, а частина – потомство з розщепленням ознаки подібно до гібридів F2: 3 домінантних до 1 рецесивної.
Заслуга Г. Менделя в тому, що він зрозумів: такі співвідношення ознак у потомстві можуть бути тільки наслідком існування відокремлених та незмінних одиниць спадковості, які передаються від покоління до покоління через статеві клітини. Для домінантного і рецесивного чинників Г. Мендель увів буквені позначення: домінантні позначалися великими літерами латинської абетки, а рецесивні – маленькими. Наприклад: А – квітки забарвлені, а – квітки білі; В – насіння гладеньке, b – насіння зморшкувате тощо.
Внаслідок ретельного аналізу одержаних експериментальних результатів Г. Мендель дійшов таких висновків.
- Оскільки вихідні сорти гороху є чистими (дають одноманітне потомство), це означає, що сорт з домінантною ознакою повинен мати два домінантні фактора (АА), а сорт з рецесивною ознакою – два рецесивні (аа).
- Статеві клітини (гамети) містять лише один фактор – домінантний (А) або рецесивний (а).
- Рослини гібридів першого покоління (F1) містять по одному фактору, одержаному через гамети від кожного з батьків, тобто А і а (Аа).
- У поколінні F1 спадкові фактори не змішуються, а залишаються відокремленими.
- Один із спадкових факторів домінує над іншим.
- Гібриди F1 утворюють два типи гамет порівну: одні з них містять фактор А, інші – фактор а.
- У процесі запліднення жіноча гамета типу А матиме рівні шанси з’єднатися як з чоловічою гаметою «А», так і з чоловічою гаметою «а». Те ж саме справедливе і для жіночих гамет типу а.
Ці висновки можна наочно представити у вигляді схеми (мал. 2).
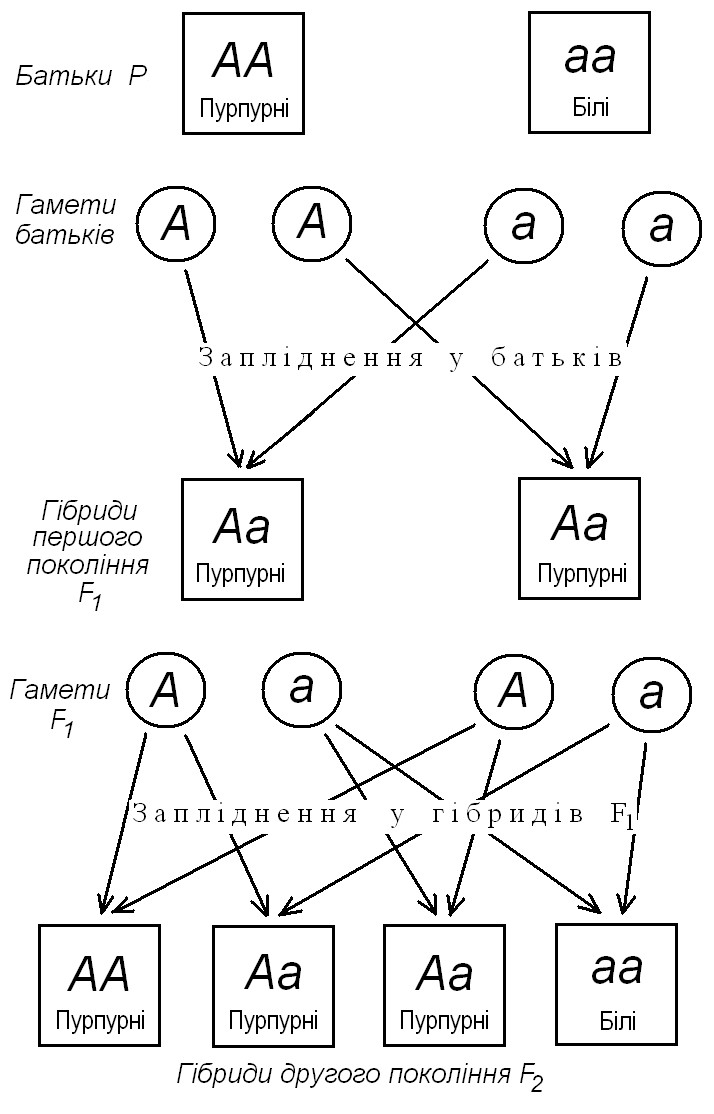
Мал. 2. Моногібридне схрещування у гороху
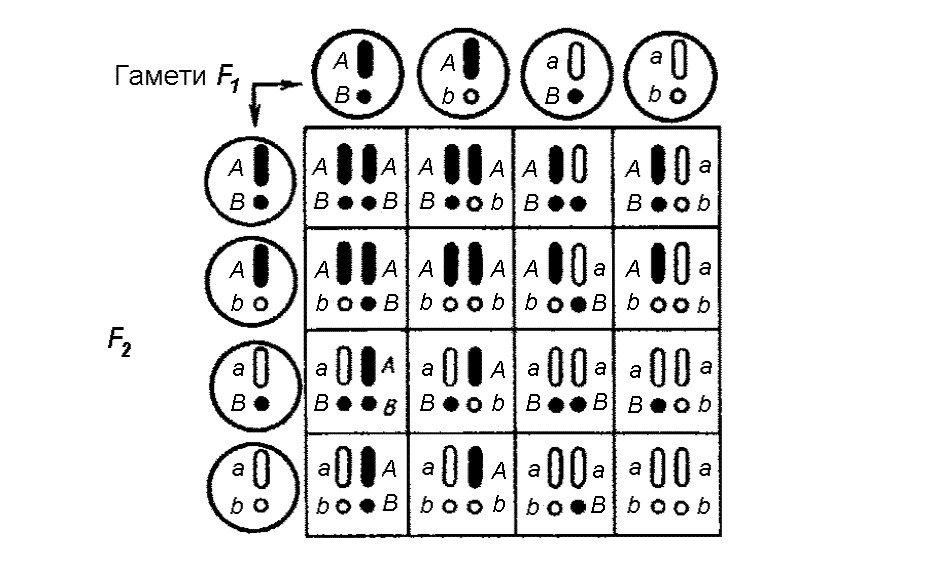
З таблиці видно, що можливі 4 різні комбінації, з яких лише одна не містить домінантний фактор, тому в цьому випадку спостерігатиметься рецесивна ознака. Три комбінації, що залишилися, представляють рослини з домінантною ознакою. Одна частина цих рослин (АА) дасть стійке потомство, яке, не розщеплюється, а дві частини, що залишилися, знову дадуть розщеплювання в співвідношенні 3 : 1 (див. також мал. 1).
У своїй публікації Г. Мендель не сформулював ніяких законів, які нині широко відомі під назвою законів Г. Менделя. За нього це зробили інші дослідники, які повторно відкрили менделеві закономірності.
Перший закон Менделя, або закон одноманітності стверджує, що при схрещуванні особин з альтернативними ознаками у гібридів першого покоління проявляються тільки домінантні ознаки.
Другий закон Менделя, або закон розщеплення, показує, що при самозапиленні гібридів першого покоління серед нащадків відбувається розщеплення ознак у співвідношенні 3 : 1, тобто утворюється 3 частини рослин з домінантною ознакою та 1 частина – з рецесивною.
Схрещування, коли вивчається лише одна пара альтернативних ознак, дістало назву моногібридного.
Дигібридне схрещування та третій закон Менделя. Після виявлення закономірностей розщеплення за однією парою альтернативних ознак Г. Мендель перейшов до вивчення успадкування двох пар таких ознак.
Схрещування особин, що розрізняються за двома парами альтернативних ознак (наприклад, насіння гладеньке жовте та зморшкувате зелене), називається дигібридним схрещуванням.
У дослідах Г. Менделя одна батьківська рослина несла домінантні ознаки (гладеньке жовте насіння), а друга – рецесивні (зморшкувате зелене насіння). Як і очікувалося, усі гібридні рослини першого покоління (F1) мали домінантні ознаки – гладеньке жовте насіння. Друге ж покоління (F2) складалося з рослин, які мали різне поєднання ознак за таким співвідношенням:
9 частин рослин з гладеньким жовтим насінням,
3 частини із зморшкуватим жовтим насінням,
3 частини з гладеньким зеленим насінням, 1 частина із зморшкуватим зеленим насінням, тобто 9 : 3 : 3 : 1.
Отже, у другому поколінні гібридів з’явилося два нові поєднання ознак насіння: зморшкувате жовте та гладеньке зелене. На підставі цього Г. Мендель зробив висновок, що спадкові чинники батьківських рослин, які об’єдналися в поколінні F1, у наступних поколіннях розділяються і поводяться незалежно – кожна ознака з однієї пари може поєднуватися з будь-якою ознакою з іншої пари. Це відкриття Г. Менделя дістало назву третього закону Менделя, або закону незалежного розподілу.
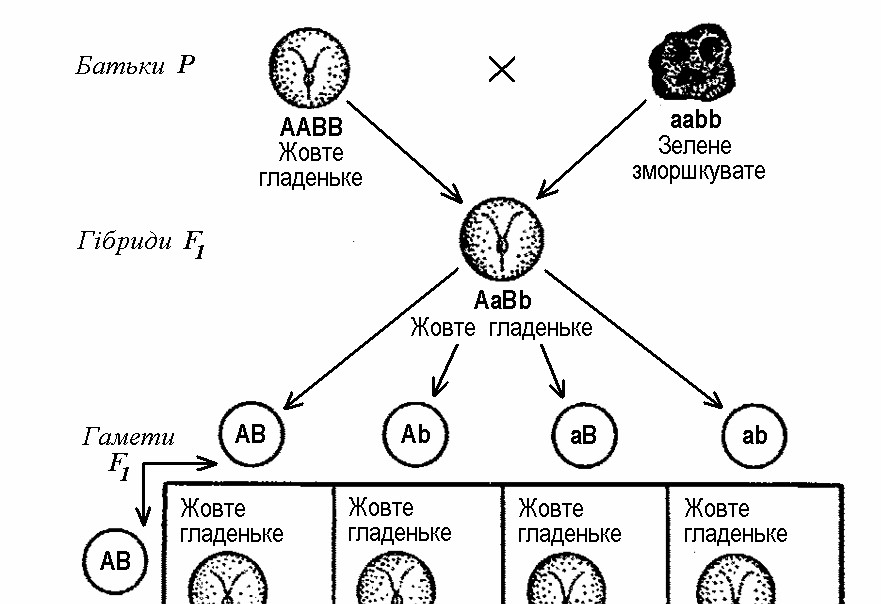
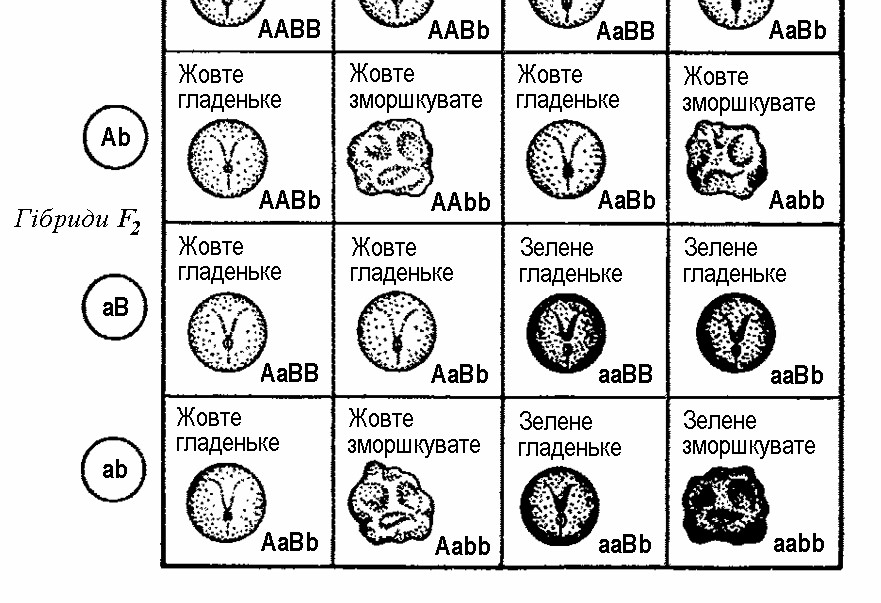
Мал. 3. Успадкування забарвлення та форми насіння у гороху
Розщеплення в умовах дигібридного схрещування можна ілюструвати за допомогою спеціальної таблиці (мал. 3), яка в генетичних дослідженнях дістала назву решітки Пеннета (в честь англійського дослідника Р. Пеннета, який її запропонував). Така таблиця забезпечує зведення помилок, які можуть виникнути при складанні всіх можливих поєднань гамет, до мінімуму. Домінантні чинники позначимо літерами А та В, а рецесивні – a та b. Тоді батьківські особини будуть мати генетичні формули ААВВ і ааbb, їхні гамети – АВ і аb, а гібриди першого покоління F1 – АаВb. Відповідно, ці гібриди можуть продукувати чотири типи гамет, після з’єднання яких формується чотири типи рослин за комбінацією ознак.
Із законів Г. Менделя можна зробити два важливих висновки:
- спадкові фактори відокремлені та незмінні; вони не змішуються, а лише перегруповуються,
- спадкові фактори схрещуваних сортів при утворенні гамет можуть утворювати нові поєднання, або рекомбінувати
Відкриття Г. Менделя не було оцінено за його життя внаслідок недостатнього рівня тодішньої науки. Лише на межі ХІХ та ХХ століть його закони були вдруге відкриті трьома .біологами, незалежно один від одного, – Г. де Фрізом, К. Коренсом та Е Чермаком, що започаткувало розвиток нової біологічної дисципліни – генетики.
2.1.2. Хромосомна теорія спадковості
Коротка історія. Ще в XVII столітті було виявлено, що організми складаються з клітин, але лише в 1831 році була відкрита найголовніша частина клітини – клітинне ядро. Згодом з’ясувалося, що ядро має досить складну будову. В 1848 році були вперше описані найважливіші його компоненти – хромосоми, проте їх особливості та функціональне значення почали активно вивчати лише в кінці XIX століття.
Хромосоми у клітинах, що не діляться, як правило, побачити неможливо. Однак під час поділу клітин їх чітко видно як тільця характерної форми (мал. 4 та 5). По мірі розвитку та вдосконалення мікроскопії стало можливим спостерігати поведінку хромосом у процесі утворення гамет і запліднення.
Наприкінці ХІХ століття було описано поведінку хромосом під час поділу клітин та виявлено, що в процесі запліднення відбувається злиття ядер чоловічої та жіночої гамет. Пізніше, після повторного відкриття законів Г. Менделя, була помічена вражаюча схожість між передачею менделевих факторів та поведінкою хромосом під час утворення гамет і запліднення. На підставі цих спостережень було висловлено припущення, що саме хромосоми є носіями менделевих чинників спадковості. На початку ХХ століття це припущення блискуче підтвердилося дослідами Т.Г. Моргана з колегами, які сформулювали хромосомну теорію спадковості. Згідно з цією теорією спадковість пов’язана з генами, розташованими в хромосомах клітинних ядер лінійно, утворюючи групи зчеплення.
Оскільки хромосоми є головним матеріальним носієм генетичної інформації, яка обумовлює схожість між поколіннями та біологічну мінливість організмів, необхідно знати, що відбувається з хромосомами під час поділу клітин..
Поділ клітин. В останній чверті ХІХ століття були описані процеси, що відбуваються в ядрах клітин тіла організму під час його росту, розвитку та регенерації пошкоджених тканин. Такий тип поділу клітин був названий мітозом . Мітоз забезпечує збільшення кількості соматичних клітин живого організму.
Мітоз – цей такий поділ клітинного ядра, при якому утворюються два дочірні ядра з наборами хромосом, ідентичними наборам материнської клітини. Разом з поділом ядра відбувається поділ цитоплазми на дві рівні частини та відновлення клітинної мембрани. (Мал. 4).
Процес утворення гамет супроводжується поділом клітин іншого типу, який дістав назву мейозу. Мейоз значною мірою подібний до мітозу, але його наслідки абсолютно відмінні. (Мал. 5).
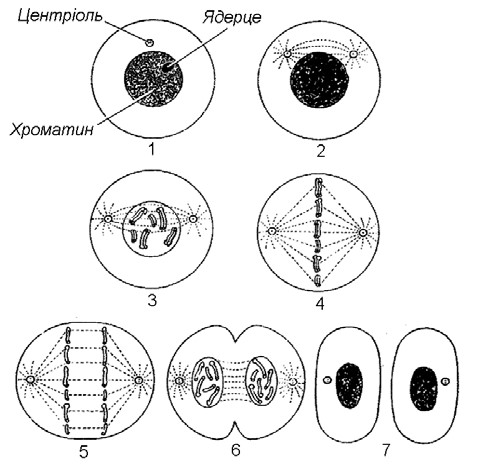
Мал. 4. Мітоз
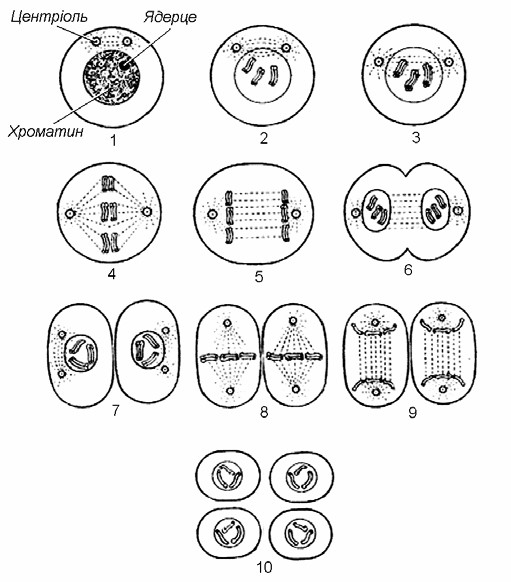
Мал. 5. Мейоз
Мейоз – це процес поділу клітинного ядра з утворенням чотирьох дочірніх ядер, кожне з яких містить удвічі менше хромосом, ніж материнське ядро. За мейозу в ядрі материнської клітини спочатку здійснюється одноразове подвоєння хромосом, після чого відбувається два послідовних поділи хромосом – перший поділ мейозу (мейоз І) та другий поділ мейозу (мейоз ІІ). Протягом мейозу І із материнського ядра утворюється два дочірні ядра, кожне з яких несе вдвічі менше хромосом порівняно з материнським ядром. Мейоз ІІ подібний до мітозу. Таким чином, за мейозу ядро ділиться двічі, а хромосоми подвоюються тільки один раз. В результаті такого поділу утворюється чотири клітини, в яких кількість хромосом вдвічі менша, ніж у материнській. З таких клітин формуються гамети.
Мейоз забезпечує збереження у наступних поколіннях видів із статевим розмноженням постійного числа хромосом у ядрах клітин тіла.
Хромосоми людини. Життя будь-якого організму, для якого характерний статевий спосіб розмноження, у тому числі й організму людини, починається з моменту злиття материнської та батьківської гамет. У яйцеклітині стільки ж хромосом, як і в сперматозооні, і вони такі ж за формою та розмірами, не дивлячись на значну різницю між чоловічою та жіночою гаметами. Число хромосом у гаметах в два рази менше, ніж в клітинах тіла. У клітинах тіла людини 46 хромосом (23 пари хромосом) – це диплоїдний набір хромосом. Відповідно, в статевих клітинах їх удвічі менше, тобто 23 – це гаплоїдний набір хромосом.
У тих клітинах, з яких будуть утворюватися гамети, можна спостерігати хромосоми, згруповані в пари. Хромосоми, що становлять пару, називають гомологічними . Вони схожі за будовою і містять спадковий матеріал, що визначає одні й ті ж ознаки. Винятком є лише статеві хромосоми, які у жінок парні та подібні за будовою – Х-хромосоми. У чоловіків же – тільки одна Ххромосома. Пару їй складає Y-хромосома, яка разюче відмінна за будовою та функціями від Х-хромосоми. (Мал. 6).
Хромосоми людини згруповані за розмірами та формою, і кожній з них присвоєно відповідний номер. Таким чином, хромосомний набір, або каріотип людини складається з 22 пар аутосом, нумерація яких зростає по мірі зменшення їх довжини, та 1 пари статевих хромосом. Графічне зображення каріотипу називається ідіограмою (Мал. 7).


Мал. 6. Хромосоми чоловіка
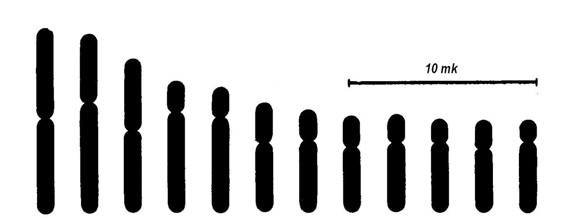
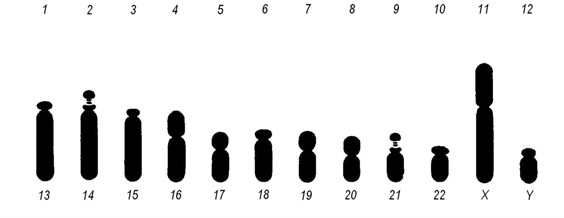
Мал. 7. Ідіограма хромосом чоловіка
Рекомбінація хромосом у процесі утворення статевих клітин. Одна із гомологічних хромосом одержана дитиною від батька, а інша – від матері. При утворенні гамет у її організмі гомологічні хромосоми спочатку кон’югують (з’єднуються) у пари, а потім розходяться до різних гамет. Дуже важливо, що хромосоми кожної пари здійснюють цей процес незалежно від інших пар. У результаті хромосоми, одержані від батька і матері, перерозподіляються по гаметах, які продукує їх нащадок, цілком випадково. При цьому в гаметах утворюються нові поєднання хромосом, відмінні від тих, що існували в батьківських гаметах, злиття яких дало життя даній людині. Така рекомбінація хромосом веде і до рекомбінації генів, що знаходяться в них, а отже, до виникнення нових поєднань ознак і збільшення генетичної різноманітності. (Мал. 8).
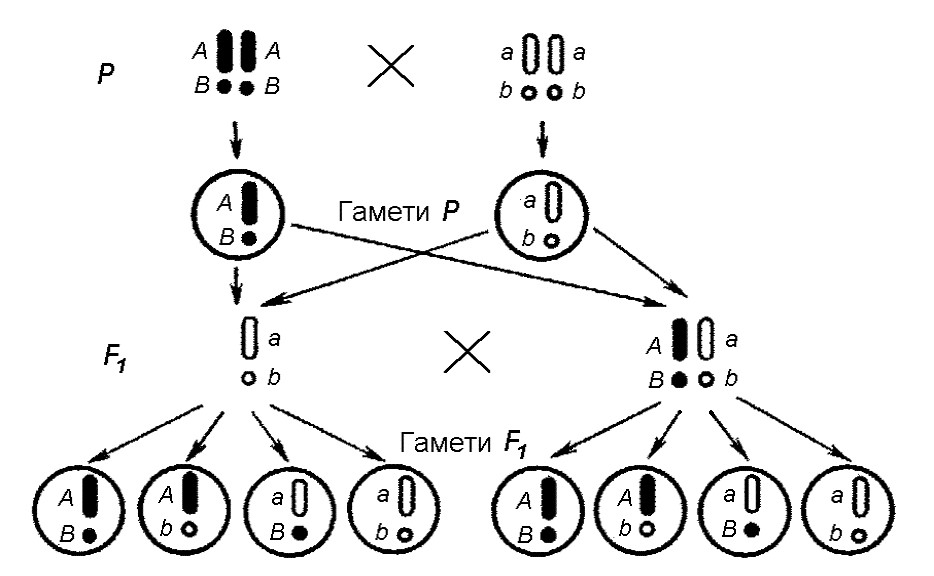
Мал. 8. Рекомбінація хромосом
Зверніть увагу на те, що рекомбінація хромосом при утворенні гамет нагадує рекомбінацію ознак за дигібридного схрещування у дослідах Г. Менделя. Г. Менделю справді повезло, що ознаки, які він вибрав для вивчення, знаходились у різних хромосомах. Якби ознаки визначалися генами, розташованими в одній хромосомі, вони б передавалися разом, і принцип незалежного успадкування ознак не проявився б.
Зчеплення та кросинговер. Якщо гени, що визначають ряд ознак, розташовані в одній хромосомі, їх називають зчепленими. Проте не слід думати, що зчеплені гени назавжди пов’язані один з одним. Насправді природа передбачила механізм, що дозволяє цим генам іноді рекомбінувати, правда, якщо вони не дуже близько один до одного розташовані в хромосомі.
Під час мейозу в процесі кон’югації гомологічних хромосом створюються умови для виникнення події, яка дістала назву кросинговеру. Кросинговер полягає в обміні ділянками між гомологічними хромосомами. (Мал. 9).
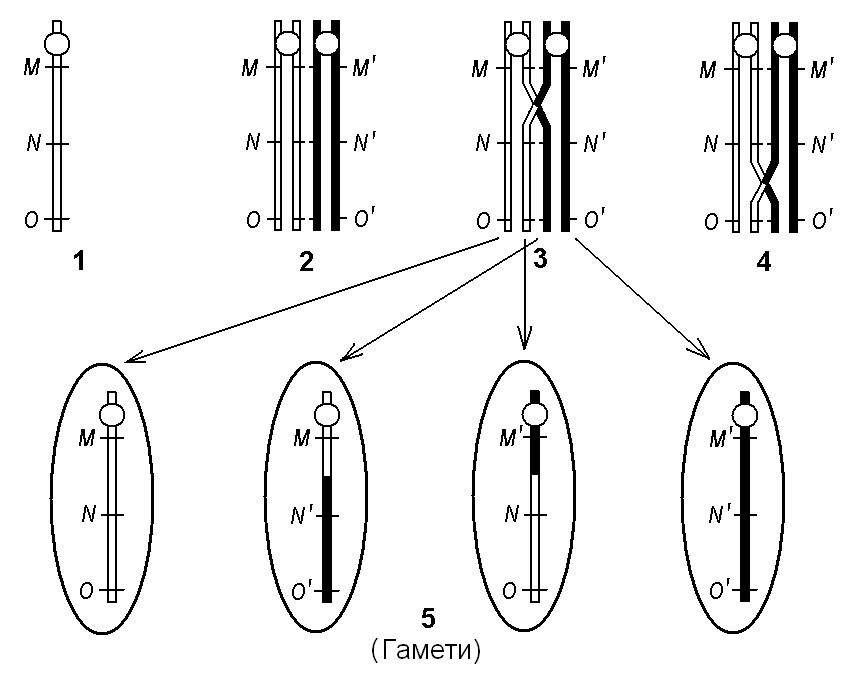
Мал. 9. Рекомбінація генів у процесі кросинговеру
Зрозуміло, що гени, достатньо далеко розташовані один від одного, повинні рекомбінувати з більшою частотою, ніж гени, розташовані близько.
Генетична унікальність індивіда. Кросинговер, поряд з незалежною рекомбінацією хромосом під час мейозу, ще більше сприяє рекомбінації генів, тим самим збільшуючи генетичну різноманітність потомства. У свою чергу, гени матері та батька лише тимчасово поєднуються в організмі нащадка, щоб потім знову розійтися в його гаметах і дати нові поєднання генів у його дітях і т.д.
Рекомбінація є основним джерелом генетичної мінливості живих організмів. Це явище настільки значне, що серед людей практично немає двох генетично однакових особин за виключенням, звичайно, справжніх (монозиготних, або однояйцевих) близнюків, які несуть однаковий набір генів. Учені підрахували, що число можливих поєднань генів людини складає 3·1047, тоді як кількість людей, які жили на Землі за всю історію людства складає 7·1010, тобто на десятки порядків менше. Тому на земній кулі практично ніколи не було, немає і не може бути бодай двох людей з однаковим набором генів.
2.1.3. Основи молекулярної генетики
Матеріальний носій спадковості. Для того, щоб здійснювалися описані вище процеси, хромосомна речовина повинна мати дві властивості – здатність до точного подвоєння та формування величезної різноманітності генів, які існують у природі.
У середині ХХ століття було доведено, що носієм генетичної інформації є дезоксирибонуклеїнова кислота (ДНК) – органічна сполука, яка разом з білком складає тіло хромосоми.
ДНК має ланцюжкову молекулярну будову, що забезпечує здатність до подвоєння та до утворення безлічі типів сполучень її елементарних одиниць – нуклеотидів. Кожний нуклеотид складається з трьох частин: азотистої основи (О), вуглеводного компоненту (дезоксирибози – Д) та залишку фосфорної кислоти (Ф). (Мал. 10).
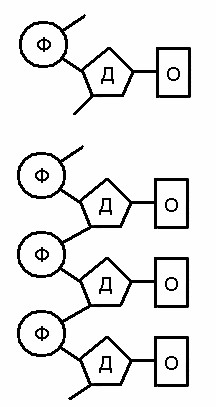
Мал. 10. Будова окремого нуклеотиду (вгорі) та фрагменту одинарного ланцюжка ДНК
У ланцюжку ДНК окремі нуклеотиди сполучені один з одним через фосфорну кислоту міцним хімічним зв’язком. Вуглеводний та фосфорний компоненти у всіх нуклеотидів однакові, але основ є чотири типи: аденін, цитозин, гуанін та тимін. Для запису генетичного коду їх позначають буквами А, Ц, Г і Т відповідно.
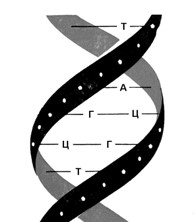
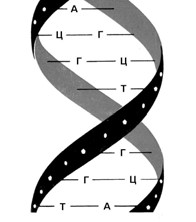
Мал. 11. Модель будови ДНК (за Ф. Уотсоном та Дж. Кріком)
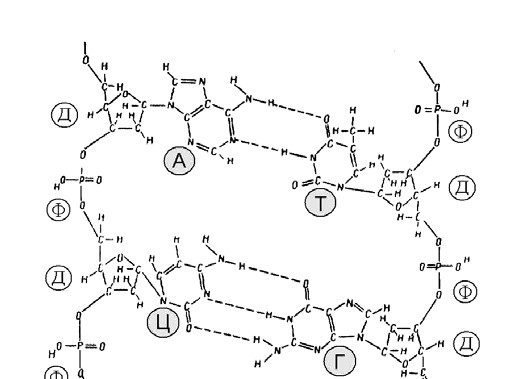
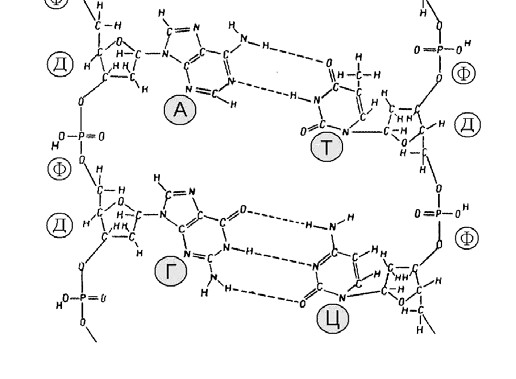
Мал. 12. Хімічна будова молекули ДНК:
А – аденін, Т – тимін, Г – гуанін, Ц – цитозин,
Ф – залишок фосфорної кислоти, Д – дезоксирибоза
Молекула ДНК утворена не одним, а двома такими ланцюжками, які з’єднуються між собою слабкими водневими зв’язками через основи. Основи пари підходять один до одного, як ключ до замка. При цьому аденін завжди парується з тиміном, а гуанін з цитозином. (Мал. 11 та 12).
Завдяки такій комплементарній (доповнювальній) будові ця подвійна молекула здатна точно відтворювати себе, утворюючи ідентичні подвійні молекули. Перед подвоєнням ДНК слабкі водневі зв’язки між основами рвуться і дві напівмолекули розходяться, як застібка-блискавка. Після цього на кожній з них добудовується нова комплементарна половинка, внаслідок чого утворюється дві нові молекули ДНК, абсолютно ідентичні початковій. Одна з них має стару «праву» сторону і нову «ліву», а інша, навпаки, – стару «ліву» та нову «праву». (Мал. 13).
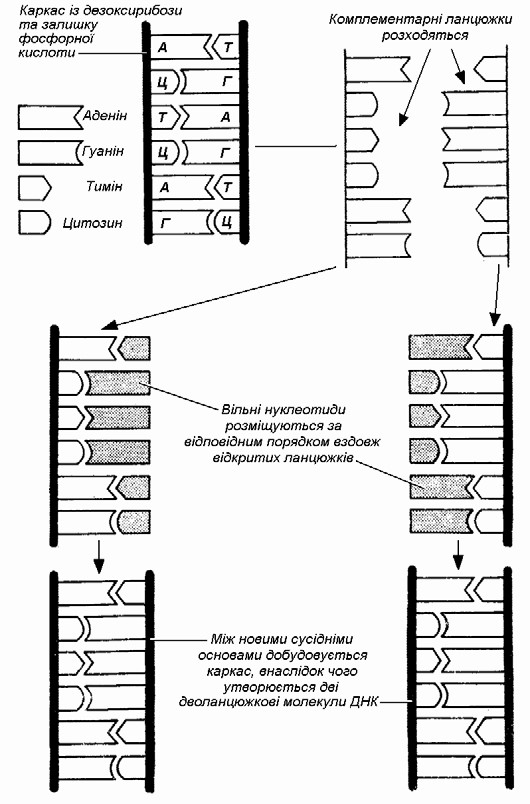
Мал. 13. Подвоєння ДНК (за Ф. Уотсоном та Дж. Кріком)
Потрібно пам’ятати, що це лише модель і що в дійсності цей процес відбувається набагато складніше.
Оскільки основи в молекулі ДНК розташовані лінійно (одна за одною) число різних комбінацій їх взаємного розташування практично необмежене, хоча основ усього чотири. Наприклад, якщо один ген містить 500 основ, то число можливих способів їх розташування складе 4500. Така комбінативна властивість забезпечує існування великої кількості різноманітних генів.
ДНК знаходиться в хромосомах разом з білками (гістоновими та негістоновими) та невеликою кількістю РНК. У кожній хромосомі міститься лише одна молекула ДНК. Під час поділу клітини хромосоми значно укорочуються, потовщуються і стають видимими під мікроскопом. Це відбувається внаслідок багаторівневої спіралізації молекули ДНК. (Мал. 14).
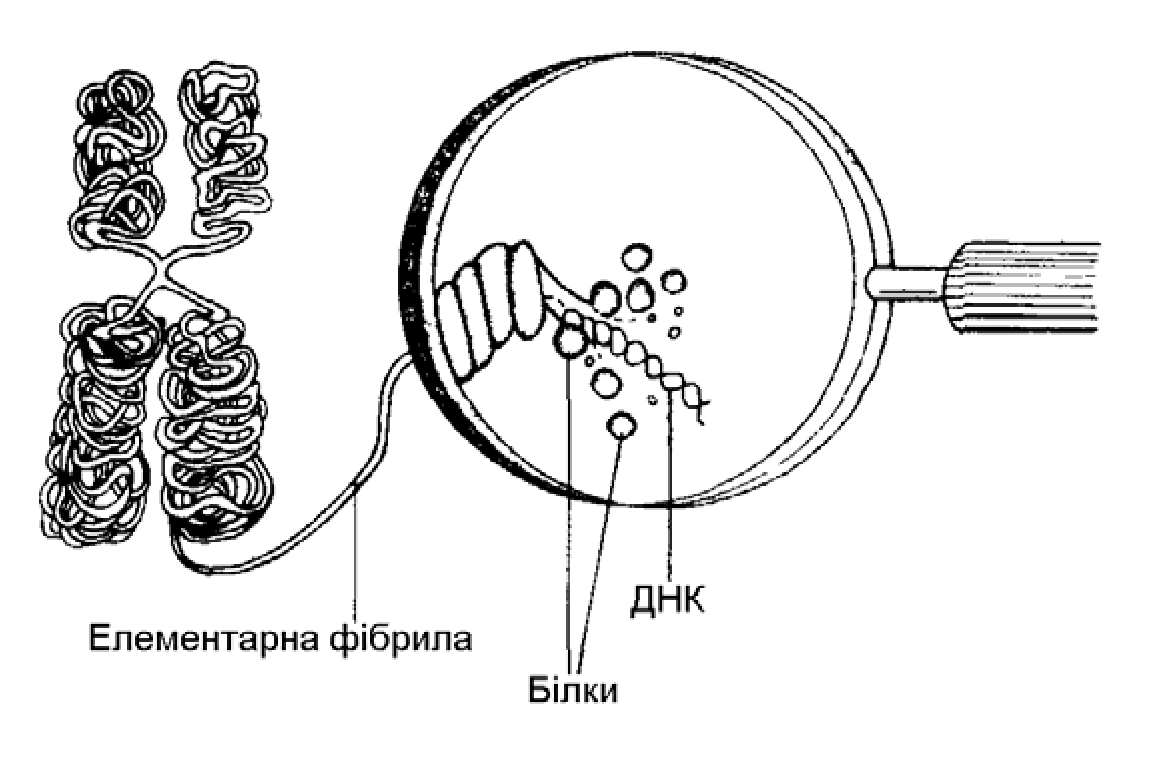
Мал. 14. Схематична будова хромосоми
Ген та його основна функція. Сучасній генетиці багато відомо про будову хромосом, про структуру та функції ДНК, але вона все ще неспроможна дати точне визначення гена. Найбільш розповсюджене сучасне уявлення стверджує, що ген – це невелика ділянка хромосоми, що виконує певну біохімічну функцію та здійснює специфічний вплив на структурні, фізіологічні та біохімічні властивості організму. Біохімічна функція гена полягає у тому, що він обумовлює синтез певного ферменту. Ферменти – це особливі білки, які в живих клітинах відіграють роль біологічних каталізаторів. За допомогою ферментів здійснюються всі біохімічні реакції обміну речовин та енергії в живих організмах. Виявилося, що гени містять інформацію про послідовність амінокислот у молекулах «первинних білків» – поліпептидів. Поліпептиди являють собою ланцюжки із амінокислот, кількість яких варіює від 6 до кількох десятків. Із певної кількості відповідних поліпептидів за допомогою особливих ферментів синтетаз утворюється молекула того чи іншого біка. Крім того, окремі поліпептиди в організмі можуть виконувати функції гормонів, біологічно активних речовин, антибіотиків тощо.
Генетичний код. Відомо, що до складу білка входить 20 різних амінокислот, а типів основ ДНК – всього 4. Яким же чином інформація про послідовність основ у молекулі ДНК перетворюється в послідовність амінокислот у молекулі білка? Виявилося, що цю умову найкраще задовольняє кодування однієї амінокислоти трьома основами. Функціональна генетична одиниця із трьох основ дістала назву триплету, або кодона. І справді, коли б одна амінокислота кодувалася двома основами, то це б дало змогу включити в молекулу білка всього тільки 16 різних амінокислот із 20. Залежність порядку розташування амінокислот у молекулах поліпептидів від порядку розташування триплетів основ у молекулі ДНК називається генетичним кодом.
(Табл 1).
Табл. 1. Генетичний код інформаційної РНК
Друга основа | |||||||||||||
У | Ц | А | Г | ||||||||||
Перша основа | У | УУУ | Фен | УЦУ УЦЦ | Сер | УАУ | Тир | УГУ УГЦ | Цис | У Ц | Третя основа | ||
УУЦ | УАЦ | ||||||||||||
УУА | Лей | УЦА УЦГ | УАА | Терм | УГА | Терм | А | ||||||
УУГ | УАГ | УГГ | Три | Г | |||||||||
ЦУУ | ЦЦУ ЦЦЦ | ЦАУ | Гіс | ЦГУ ЦГЦ | У Ц | ||||||||
ЦУЦ | ЦАЦ | ||||||||||||
Ц | ЦУА | Лей | ЦЦА ЦЦГ | Про | ЦАА | Глу | ЦГА ЦГГ | Арг | А Г | ||||
ЦУГ | ЦАГ | ||||||||||||
А | АУУ | ІЛей | АЦУ АЦЦ АЦА | Тре | ААУ | Асн | АГУ АГЦ | Сер | У Ц | ||||
АУЦ | ААЦ | ||||||||||||
АУА | ААА | Ліз | АГА АГГ | Арг | А Г | ||||||||
АУГ | Мет | АЦГ | ААГ | ||||||||||
Г | ГУУ | Вал | ГЦУ ГЦЦ ГЦА ГЦГ | Ала | ГАУ | Асп | ГГУ ГГЦ | Глі | У Ц А Г | ||||
ГУЦ | ГАЦ | ||||||||||||
ГУА | ГАА | Глу | ГГА ГГГ | ||||||||||
ГУГ | ГАГ | ||||||||||||
Примітка. Генетичний код ДНК містить комплементарні основи, а У замінений у ньому на Т.
Терм – термінатор (стоп-кодон); основи: А – аденін, Г – гуанін, Т – тимін, Ц – цитозин, У – урацил; амінокислоти: Ала – аланін, Арг – аргінін, Асн – аспарагін, Асп – аспарагінова кислота, Вал – валін, Гіс – гістидин, Глі – гліцин, Глу – глютамін, ІЛей – ізолейцин, Лей – лейцин, Ліз – лізин, Мет – метіонін, Про – пролін, Сер – серин, Тир – тирозин, Тре – треонін, Три – триптофан, Фен – фенілаланін, Цис – цистеїн.
61 кодон визначає відповідні амінокислоти, і всі амінокислоти, за виключенням триптофану та метіоніну, кодуються кількома кодонами. Кодони-синоніми звичайно утворюють групи, в яких дві перші основи в кодоні є загальними, а третя – варіює.
Генетичний код є універсальним, бо в усіх живих організмів одні й ті ж амінокислоти кодуються одними й тими ж триплетами. Цікаво, що певна амінокислота може кодуватися більш ніж одним триплетом. Згадайте, що число можливих триплетів 64, а число амінокислот 20. Крім того, код не перекривається, тобто кожна основа може належати тільки одному триплету
Механізм синтезу білків (поліпептидів) у клітині надзвичайно складний. Він вимагає участі іншого виду нуклеїнових кислот – рибонуклеїнової кислоти (РНК) та особливих клітинних органел – рибосом. (Мал. 15).
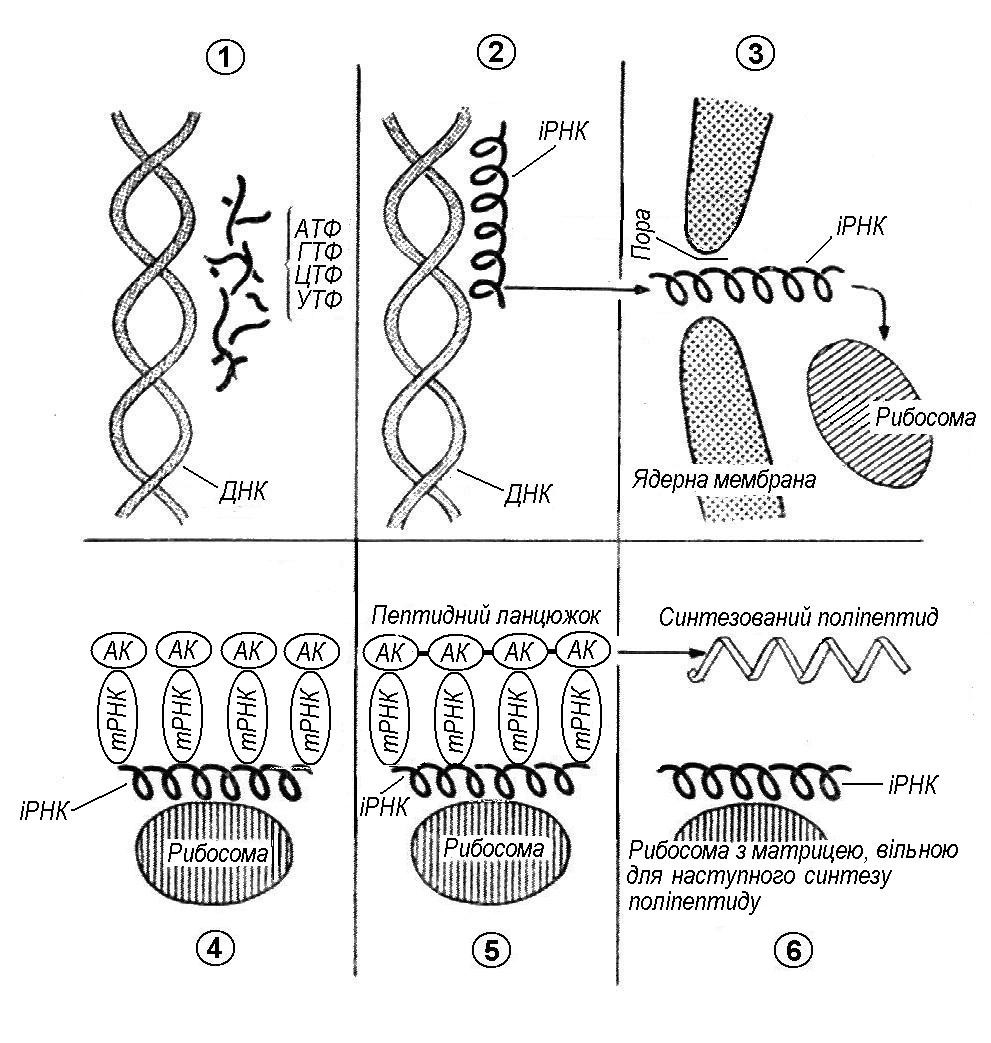
Мал. 15. Схема біосинтезу поліпептиду
Сучасні молекулярно-генетичні дослідження показали, що будова гена та принцип зчитування інформації для синтезу білка у еукаріотів (організмів, клітини яких мають справжнє ядро) відрізняється від такого у прокаріотів (одноклітинних організмів, які позбавлені справжнього ядра). Виявилося, що гени еукаріотів містять як кодовані ділянки, які несуть інформацію для синтезу специфічного білка (вони були названі екзонами), так і некодовані (названі інтронами). Причому, некодованих ділянок може бути в декілька разів більше, ніж кодованих. А у людини з усієї генетичної ДНК лише близько 5% складають кодовані ділянки.
Завдяки такій будові один і той же ген еукаріотів може нести інформацію для кодування не одного поліпептиду, як у прокаріотів, а, залежно від специфіки тканини, в якій вони функціонують, – великої кількості різних поліпептидів. Наприклад, деякі гени можуть нести код для синтезу майже 40 тис. поліпептидів. Це досягається за рахунок зміни порядку зчитування кодованих ділянок гена. Цікаво, що у людини, за сучасними даними, 74% генів працюють саме за таким принципом.
2.1.4. Гени в хромосомах
Як відомо з хромосомної теорії спадковості, гени в хромосомах розташовуються лінійно. Місце в хромосомі, де знаходиться даний ген, називається локусом цього гена. Певний локус може займати лише одна з форм одного й того ж гена – домінантна, рецесивна або інша. Такі різні стани гена називаються алелями . Для більшості генів відомі лише два алелі – домінантний та рецесивний. Але часто має місце так званий множинний алелізм, коли існує ціла низка станів певного гена.
Найпростішим прикладом множинного алелізму може бути успадкування груп крові у людини за системою АВ0. Кожна людина має одну з чотирьох груп крові, які зумовлені взаємодією трьох множинних алелей одного й того ж гена – IА, IВ та I0. Алелі IА та IВ є домінантними, а I0 – рецесивний.
Нижче подані групи крові та сполучення пар алелей, які їх визначають.
І, або 0 – І0 І0;
ІІ, або А – ІА ІА, ІА І0; ІІІ, або В – ІВ ІВ, ІВ І0; IV, або АВ – ІА ІВ.
У випадку IV групи крові домінування того чи іншого алеля не спостерігається, але відсутній і проміжний ефект. Кров цієї групи одночасно проявляє ознаки ІІ та ІІІ групи.
Алелі гена, розташовані в тотожних локусах гомологічних хромосом, можуть бути однаковими – домінантними (АА) або рецесивними (аа) Таке сполучення пари алелей одного гена називається гомозиготним. Якщо ген представлений двома різними алелями (Аа), то такий стан буде гетерозиготним.
2.1.5. Мутації
Генні мутації. Хоча реплікація (подвоєння) ДНК під час мейозу або мітозу відбуваються з досить великою точністю, час від часу трапляються помилки, які призводять до зміни послідовності нуклеотидів у ланцюжку ДНК. Таке явище називається мутацією гена. Мутація може бути спричинена заміною однієї пари основ на іншу, а також втратою нуклеотидів чи появою одного або кількох нових нуклеотидів. Більш суттєві наслідки будуть у випадку втрати або появи нуклеотидів. Тоді порядок прочитання триплетів неминуче зсовується на одну-дві основи вправо або вліво, внаслідок чого генетичний код зчитується неправильно. Якщо випали чи добавились одночасно три основи, то зміни торкнуться лише однієї амінокислоти, а решта ланцюжка залишиться нормальною.
Генні мутації, що виникають у гаметах, справляють на організм різноманітний вплив. Більшість із них є летальними (смертельними), оскільки викликають дуже серйозні порушення розвитку. Відомо, наприклад, що у людини близько 20% вагітностей закінчуються природним викиднем протягом 12 тижнів з моменту зачаття, і близько половини цих випадків спричинені генними мутаціями..
Переважна більшість генних мутацій рецесивна по відношенню до “нормального” алеля. Майже всі вони шкідливі для організму, бо викликають функціональні та структурні порушення, що призводять до захворювання, яке може закінчитися смертю. Рецесивна мутація проявиться лише тоді, коли в геномі організму два мутантних алеля гена складуть гомологічну пару. Інколи виникають і домінантні мутантні алелі, які проявляються відразу. Домінантні мутації теж можуть бути шкідливими.
Мутації, що відбуваються в соматичних клітинах, передаються лише тим клітинам, які утворюються з клітини-мутанта шляхом мітозу. Вони можуть проявитися лише в тому організмові, де вони виникли, і потомству не передаються.
Хромосомні аберації. Під час мейозу в процесі кон’югації гомологічних хромосом може трапитися порушення нормальної рекомбінації, внаслідок чого з’являються гамети з пошкодженими хромосомами. Такі пошкодження називаються хромосомними абераціями. Відома ціла низка хромосомних аберацій – інверсії, делеції, дуплікації, транслокації та інші. Інверсією називається обертання окремої ділянки хромосоми на 1800. Втрата певної ділянки хромосоми – це делеція , а подвоєння – дуплікація . Інколи має місце обмін ділянками між негомологічними хромосомами. Така хромосомна аберація дістала назву транслокації .
Окрім перебудов у межах окремих хромосом, трапляються випадки нерозходження окремих хромосом під час мейозу. Це призводить до того, що в якійсь гаметі може не вистачати окремої хромосоми або бути зайва. Більшість таких порушень веде до загибелі плоду ще на ранніх стадіях розвитку. Проте інколи при цьому народжуються хоча і життєздатні особини, але з тяжкими порушеннями фізичного та психічного розвитку.
Хромосомні аберації, як і генні мутації, можуть призводити до різноманітних порушень розвитку організму аж до летального кінця.
2.1.6. Генетична рівновага в популяціях
У попередньому матеріалі йшла мова про закономірності успадкування якісних ознак у межах кількох поколінь однієї пари батьківських особин. Тепер нашим об’єктом аналізу буде популяція. Популяція – це сукупність особин одного виду, яка тривалий час займає певний простір та відтворює себе протягом значної кількості поколінь.
Для усвідомлення генетичних процесів у популяції, які визначаються однією парою алелей, розглянемо просту модель успадкування груп крові людини за системою MN. Група крові за цією системою визначається двома алелями LМ та LN, які успадковуються незалежно від алелей I за системою AB0. Гомозиготи LМLМ мають групу крові М, гомозиготи LNLN – групу крові N, а гетерозиготи LМLN – групу крові МN.
Уявимо собі замкнуту людську популяцію, в якій є певна кількість алелей LМ та LN. Частота наявності кожного з цих алелей може мати різні значення в межах від 0 до 100%. В одному випадку вся популяція може бути представлена тільки гомозиготами LМLМ, у іншому – гомозиготами LNLN. Не виключено, що алелі LМ та LN у популяції представлені порівну, і частота кожного з них буде дорівнювати 50%, або 0,5.
Може також бути, що в популяції представлені всі три типи поєднань алелей, а частота наявності кожного алеля складає 0,5 (50%). Зрозуміло, що в такій популяції з рівною вірогідністю будуть утворюватися гамети з алелем LМ (0,5) та гамети з алелем LN (0,5). Якщо шлюби в популяції відбуваються випадково, то ми можемо скористатися решіткою Пеннета, щоб узнати частоту утворення гомозигот і гетерозигот у наступному поколінні (значення частот перемножуються). (Табл. 2).
Табл. 2. Частоти алелей груп крові за системою MN у популяції
Жіночі гамети | |||
0,5 LM | 0,5 LN | ||
Чоловічі гамети | 0,5 LM | 0,25 LMLM | 0,25 LMLN |
0,5 LN | 0,25 LMLN | 0,25 LNLN | |
Виявилося, що частоти наявності гомозигот (LMLM та LNLN) у нашій популяції складають по 0,25, тоді як частота наявності гетерозигот (LMLN) у два рази вище – 0,5. Частоти ж наявності кожного алеля (LМ та LN) будуть, як і до цього, однаковими – по 0,5. Відповідно в наступному поколінні цієї популяції (F2), за умови збереження випадковості шлюбів, будуть знову одержані ті ж співвідношення.
Звичайно, в популяціях спостерігаються самі різні частоти наявності алелей. До того ж між алелями гена можуть існувати домінантно-рецесивні відношення і тоді гетерозиготи за зовнішнім проявом ознаки можуть співпадати з домінантними гомозиготами. За таких обставин частоти наявності самої ознаки не співпадатимуть з частотами наявності гомозигот і гетерозигот.
Розглянемо це на моделі людській популяції з успадкуванням форми волосся. Кучеряве волосся є домінантною ознакою, а пряме – рецесивною. Позначимо алель, яка визначає кучеряве волосся літерою H, а алель прямого волосся – h. Тоді гомозиготні особини за домінантним алелем (HH) та гетерозиготні особини (Hh) матимуть кучеряве волосся, а гомозиготні особини за рецесивним алелем (hh) – пряме.
Нехай аналізована нами популяція є ізольованою, шлюби в ній відбуваються випадково, а частоти наявності алелей H та h відповідно складають 0,1 та 0,9. Звичайно частоти гамет, які несуть ці алелі, будуть відповідно такими ж. Знову за допомогою решітки Пеннета визначимо частоту різних поєднань алелей в поколінні F1. (Табл. 3).
Табл. 3. Частоти алелей форми волосся у популяції
Жіночі гамети | |||
0,1 H | 0,9 h | ||
Чоловічі гамети | 0,1 H | 0,01 HH | 0,09 Hh |
0,9 h | 0,09 Hh | 0,81 hh | |
Таким чином, у поколінні F1 частоти гомозигот і гетерозигот будуть наступними:
HH –– 0,12 = 0,01; Hh –– 2 · 0,1 · 0,9 = 0,18; hh –– 0,92 = 0,81.
Зверніть увагу на те, що сума цих частот складає одиницю (0,01 + 0,18 + 0,81). Якщо визначити частоту наявності самої ознаки, то виявиться, що в даній популяції кількість людей з кучерявим волоссям складає 19%, а з прямим – 81%.
Тепер поцікавимося, які гамети буде продукувати нове покоління? Гомозиготи HH даватимуть лише гамети з алелем H, гетерозиготи Hh – вірогідно однакову кількість гамет з алелем H і алелем h, а гомозиготи hh лише гамети з алелем h. Таким чином, частоти наявності гамет з алелем H і алелем h будуть наступними:
H –– 0,01 + 1/2 · 0,18 = 0,10; h –– 0,81 + 1/2 · 0,18 = 0,90
Виходить, що співвідношення частот алелей у замкненій популяції за умови випадковості шлюбів зберігається і в наступному поколінні. Цю закономірність можна записати у вигляді буквених виразів. Коли ми позначимо частоти алелей літерами p та q, то зможемо записати, що в популяції p + q = 1. Тепер підставимо літерні позначення частот алелей в решітку Пеннета (Табл. 4).
Табл. 4. Частоти алелей форми волосся у популяції в літерних значеннях
Жіночі гамети | |||
p H | q h | ||
Чоловічі гамети | p H | p2 HH | pq Hh |
q h | pq Hh | q2 hh | |
На підставі здійснених у таблиці розрахунків можна записати, що
р2 + 2рq + q2 = 1, або (p + q)2 = 1
Ця формула відображає взаємозв’язок частот гомозигот і гетерозигот в популяції, або закон Харді–Вайнберга, який названо за іменами його авторів.
Звичайно, в дійсності таких ідеальних популяцій людей практично не існує. Всі сучасні популяції людей відкриті, внаслідок чого здійснюються чисельні шлюби з особинами інших популяцій. Крім того в будь-якій популяції має місце природний мутаційний процес. Усе це значною мірою впливає на картину розподілу частот алелей, а отже й частот гомозигот і гетерозигот у популяції. Таким чином, в реальних популяціях закон Харді–Вайнберга не буде справджуватися.
2.2. Генетика кількісних ознак
У попередньому розділі були описані закономірності успадкування якісних, або дискретних ознак, які не мають проміжних форм. Ці закономірності можна зображати у вигляді достатньо простих схем. Згадаймо, що якісні ознаки звичайно визначаються одним-двома головними генами, у яких може бути два або кілька алелей, а зовнішні умови практично не впливають на ступінь їх прояву. Таким ознаками у людини є групи крові, форма волосся (кучеряве – пряме), ямка на підборідді та багато інших, у тому числі патологій.
Ознак, які визначаються виключно генами і на які умови життєвого середовища не справляють ніякої дії, у людини дуже мало. Більшість же ознак є результатом взаємодії численних генів між собою та з факторами довкілля. Це – кількісні ознаки, які проявляються у вигляді неперервної мінливості від мінімального до максимального їх вираження. До таких ознак у людини відносяться зріст, маса, колір шкіри та волосся, інтелект, темперамент тощо.
Вивчати закономірності успадкування кількісних ознак набагато складніше, ніж якісних, бо у цьому випадку доводиться користуватися методами варіаційної статистики.
2.2.1. Визначення кількісних ознак
Звичайно кількісні ознаки вимірюються різноманітними способами в залежності від типу ознаки та мети дослідження. Зріст вимірюється спеціальною лінійкою і визначається в сантиметрах. Маса тіла визначається в кілограмах з точністю до десятої частини. Деякі ознаки, наприклад, рівень інтелект чи темперамент визначається за допомогою спеціальних тестів.
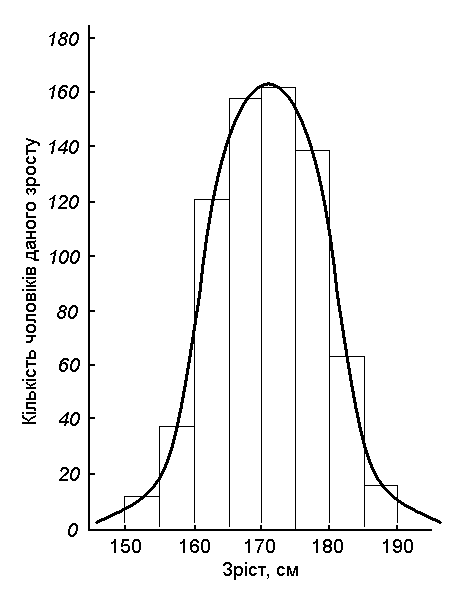
Мал. 16. Діаграма та крива розподілу частот зросту людини
В результаті таких вимірювань достатньої кількості особин (як правило, не менше тисячі) та групування їх за правилом зростання ми одержимо неперервний ряд мінливості. Цей ряд можна представити у вигляді діаграми або графіка (мал. 16).
На малюнку зображено розподіл по зросту понад 1000 дорослих чоловіків. Прямокутники показують кількість чоловіків, які мають певний зріст. Наприклад, 14 чоловіків мали зріст від 151 до 155 см, 36 чоловіків – від 156 до 160 см, 120 чоловіків – від 161 до 165 см і т.д. Ці ж дані можна представити у вигляді дзвоноподібної кривої, яка називається кривою нормального розподілу.
2.2.2. Генотип та фенотип
Для подальшого аналізу кількісних ознак необхідно познайомитися з такими поняттями як генотип і фенотип. Ми вже знаємо, що спадкова інформація про ознаки та властивості організму міститься в його генах. Уся сукупність генів даного організму називається його генотипом. Свій генотип кожна людина одержує у момент зачаття і несе його без жодних змін через усе своє життя. Протягом життя активність генів може мінятися, але їх склад залишається незмінним.
Поняття генотип не тотожний поняттю геном. Генотип притаманний певній особині, а геном – це сукупність генів, характерна для гаплоїдного набору хромосом даного виду організмів.
Фенотип являє собою сукупність усіх ознак та властивостей організму в кожний період його життя. Це і зовнішній вигляд, і внутрішня будова, і фізіологічні процеси, і особливості поведінки, спостережувані у даний момент.
Якщо генотип успадковується і залишається незмінним протягом життя індивіда, то фенотип не успадковуються – він розвиваються. Фенотип є результатом дії генотипу лише частково, бо в його становленні значну роль відіграють умови зовнішнього середовища.
Між генетиками весь час ведуться дискусії відносно того, що важливіше для розвитку організму – генетична конституція чи середовище. Особливо гострі суперечки мають місце там, де справа торкається поведінки людини та її психологічних характеристик – розумових і творчих здібностей, темпераменту тощо.
Здоровий глузд підказує, що важливі обидва фактори, бо фенотип є результатом взаємодії між генотипом і середовищем. Якщо людина не буде мати природжених здібностей, наприклад, скрипаля, то ніякі зусилля навчання та виховання не створять видатного музиканта. І навпаки, якщо людина матиме вроджені музичні здібності, то без вчасного відповідного навчання та виховання цей талант буде загублено.
2.2.3. Фактори виникнення кількісної мінливості
Тимчасово знехтуємо впливом факторів оточуючого середовища на фенотип і зосередимо свою увагу тільки на проблемі сумісної дії багатьох генів, яка сама по собі спричинює кількісну мінливість.
Закономірності успадкування кількісних ознак візуально чи за допомогою звичайних вимірювань виявити неможливо. Справа у тому, що у гібридологічних експериментах, наприклад, на рослинах усі наступні покоління мають значення ознаки (довжина стебла, розмір листків, кількість насіння тощо), проміжне між батьківськими формами. Для генетичного аналізу таких явищ користуються спеціальними математичними методами., про які мова йтиме пізніше.
Одним із основних чинників кількісної мінливості є полімерні гени.
Полімерні гени. Ще на початку ХХ століття було виявлено, що окрема кількісна ознака обумовлена сумісною дією певної сукупності генів, механізм передачі кожного з яких наступним поколінням здійснюється за законами Г. Менделя. Такі гени були названі полімерними, а ознаки, що визначаються такими генами, – полігенними.
При полімерному успадкуванні ефекти домінантних алелей можуть проявлятися двома способами.
У одних випадках інтенсивність прояву ознаки прямо пропорційна кількості домінантних алелей полімерного гена. Такий тип полімерії називається кумулятивним (накопичувальним).
Яскравим прикладом такого типу полімерії може бути успадкування кольору шкіри у людини, яке визначається 4-ма парами алелей. При цьому корінний мешканець тропічної Африки, який відрізняється найтемнішим кольором шкіри, має таку сукупність полімерних генів: P1P1P2P2P3P3P4P4 (усі алелі домінантні), а, наприклад, норвежець – p1p1p2p2p3p3p4p4 (усі алелі рецесивні). Звичайно, населення середземноморських країн у своєму генотипі має приблизно половину домінантних алелей, які визначають колір шкіри, у різноманітних комбінаціях (P1P1P2P2p3p3p4p4; P1p1P2p2P3p3P4p4 тощо). Якщо ми зобразимо результати визначення кольору шкіри достатньої вибірки людей у вигляді діаграми чи кривої, то одержимо картину, подібну до розподілу частот зросту людини. (Мал. 16)
Але зустрічаються випадки полімерії, коли достатньо наявності хоча б одного домінантного алеля, щоб ознака проявилася максимально, і збільшення кількості інших домінантних алелей помітного ефекту не справляє. При цьому у F2 має місце розщеплювання типу 15 : 1 (за умови 2-х пар алелей) або 63 : 1 (за умови 3-х пар алелей) без наявності проміжних значень ознаки.
Такий тип полімерії називається некумулятивним.
Характерним прикладом некумулятивної полімерії може бути успадкування оперення ніг у курей. Внаслідок схрещування порід, що мають оперені та неоперені ноги, усе покоління F1 матиме оперені ноги. В поколінні F2 відбувається розщеплення на два фенотипові класи за співвідношенням 15/16 з опереними ногами та 1/16 з неопереними. Таким чином, у даному випадку ознака контролюється двома парами алелей. Тоді батьківська порода курей з опереними ногами буде гомозиготною за домінантними алелями обох генів (L1L1L2L2), а з неопереними – гомозиготною за рецесивними алелями (l1l1l2l2).
Перше покоління гібридів між цими породами матиме формулу L1l1L2l2. Кури другого покоління з опереними ногами будуть мати у генотипі від 1 до 4 домінантних алелей (L1l1l2l2 ……. L1L1L2L2), а з неопереними – усі 4 алеля рецесивних (l1l1l2l2).
Умови середовища та норма реакції. Дуже часто неперервний характер може мати й мінливість, яка цілком визначається умовами середовища. Наприклад, якщо генетично ідентичні рослини (клони, чисті лінії) вирощувати в різноманітних умовах (кількість поживних речовин, води, тепла, світла тощо), то одержимо чітку неперервну мінливість за різними ознаками (висота стебла, розмір листків, кількість та маса насіння тощо). Сучасна генетика для досліджень здатна також створювати чисті, або інбредні лінії (близькоспоріднене запліднення) та клони (одержання організмів із соматичних клітин) тварин, у тому числі ссавців. Явище впливу середовища на фенотип можна спостерігати і у людини, коли справжніх (однояйцевих) близнюків виховувати в різних умовах. Як засвідчили численні спостереження, ці близнюки відрізняються між собою за фізичним, психічним та розумовим розвитком.
У розглянутих вище випадках генотип організмів є постійною величиною, а фенотип – функцією умов зовнішнього середовища. Для аналізу цього явища необхідно ввести таке генетичне поняття як норма реакції.
Якщо даний генотип за певною ознакою чутливий до змін умов середовища, то, помістивши безліч однакових генотипів у різні строго фіксовані умови середовища, ми одержимо безліч різноманітних фенотипів. На підставі вимірювання значень даної ознаки ми зможемо побудувати графік залежності характеристики фенотипу від параметрів середовища. Цей графік і буде характеризувати норму реакції даного генотипу за даною ознакою. Різні генотипи за однією і тією ж ознакою мають різні норми реакції. (Мал. 17).
На малюнку представлені результати дослідження залежності густини шерсті у двох інбредних ліній мишей від температури оточуючого середовища. Для спрощення аналізу припустимо, що ця залежність носить лінійний характер: чим вище температура, тим менше густина шерсті тварин. Дані генотипи різняться за чутливістю до температурних умов. Генотип М1 виявився менш чутливим до температури, ніж М2, про що свідчить менший нахил прямої на графіку І. Але при певній температурі фенотипи тварин, що належать до різних інбредних ліній, будуть співпадати (точка перетину В двох ліній). Таким чином, ми не зможемо однозначно виявити, як саме відрізняються ці дві інбредні лінії мишей за густиною шерсті. І справді. За низької температури середовища (ліва частина графіка) генотип М2 переважає генотип М1 за густиною шерсті. Існує також певна температура (точка В), коли за густиною шерсті генотипи не будуть відрізнятися. І нарешті, за підвищеної температури середовища (права частина графіка) за густиною шерсті буде переважати генотип М1. За даної ситуації ми з впевненістю можемо сказати лише про те, що ці дві лінії мишей за густиною шерсті мають різні норми реакції на температурні умови зовнішнього середовища.
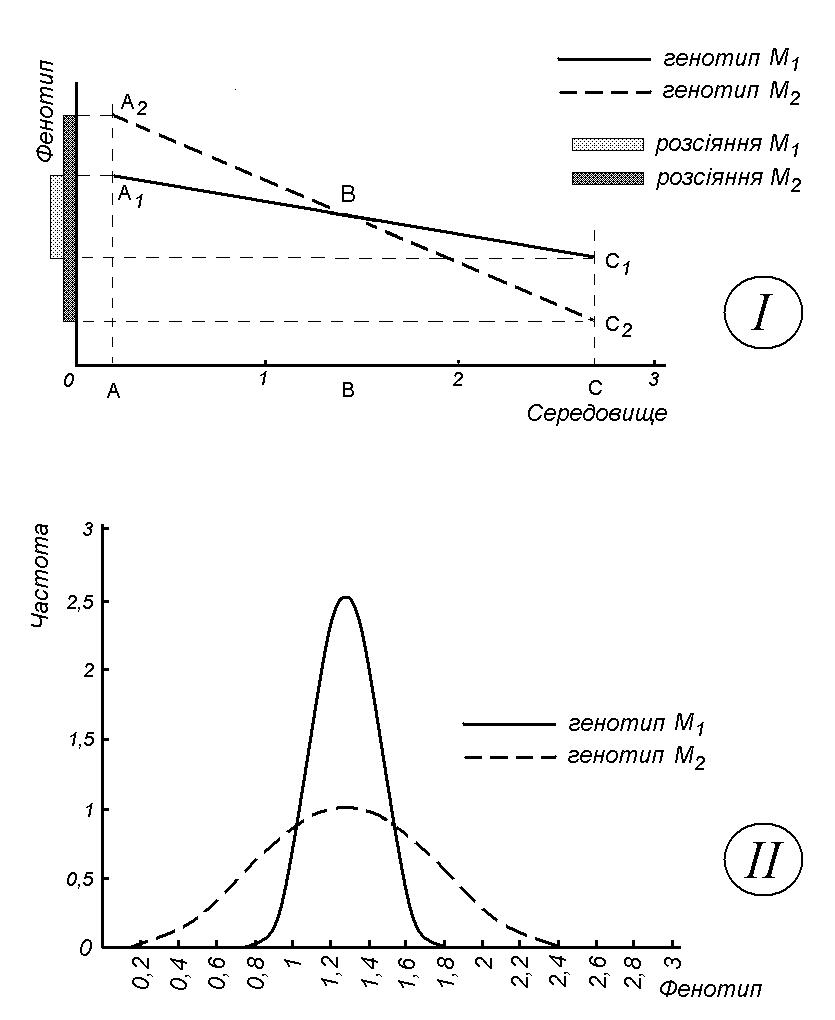
Мал. 17. Норми реакції (І) та криві розподілу частот (ІІ) двох генотипів з різною чутливістю до змін умов середовища
Якщо ми спроектуємо відрізки А1С1 (генотип М1) та А2С2 (генотип М2) на вісь фенотипу, то побачимо, що в одному й тому ж діапазоні умов середовища розсіяння фенотипної мінливості у більш чутливого до середовища генотипу М2 буде значно більшим, ніж у менш чутливого генотипу М1. Ця закономірність чітко виявляється і за кривими розподілу частот фенотипів (графік ІІ малюнка). Числовий вираз такого розсіяння у генетиці називається дисперсією. У розглянутому прикладі ми маємо справу з середовищною дисперсією. Отже, чим вища чутливість генотипу до умов середовища, тим більше його фенотипна дисперсія, яка визначається особливостями середовища існування.
Таким чином, фенотип визначається не лише генотипом. Внаслідок цього порівнювати різні генотипи потрібно не за їх фенотипами, а за нормами реакції (дисперсіями). Недаремно генетики говорять, що успадковуються не певні ознаки, а норми реакцій.
2.2.4. Співвідношення впливу генотипу та середовища
За фенотипом люди відрізняються один від одного принаймні з двох причин. По-перше, кожна людина має свій, тільки їй притаманний генотип, що приводить до виникнення генетично обумовленої мінливості. По-друге, кожна людина розвивається в особливих умовах середовища, що спричинює середовищну мінливість. Розглянемо, як із генетичної та середовищної мінливості формується загальна мінливість, або фенотип.
Уявімо собі гіпотетичну популяцію людей, у якій ми провели вимірювання якоїсь кількісної ознаки у 1000 осіб. Припустимо, що дана ознака визначається геном, який може існувати в трьох різних формах, тобто має три алеля. Позначимо їх літерами K, L, M. Тоді генотипи популяції будуть мати усі можливі комбінації алелей (табл. 5). Нехай люди з генотипом KK мають найнижчі показники ознаки, а люди з генотипом MM – найвищі. Генотип LL займає проміжне положення. Ефекти домінування відсутні, і гетерозиготи виявляються проміжними між гомозиготами.
Ми знаємо, що на конкретну величину ознаки роблять вплив не тільки гени, але й умови існування, наприклад, якість харчування, сімейний догляд у дитинстві тощо. Кожний генотип буде характеризуватися певним середнім значенням ознаки та розсіянням (дисперсією) її вимірних значень навколо середнього значення. Числове значення дисперсії розраховується за допомогою спеціальних формул варіаційної статистики. Чим вища чутливість певного генотипу до умов середовища, тим більша різноманітність його фенотипів, а отже і дисперсія. Зрозуміло, що це буде середовищна дисперсія.
Генотип | Кількість осіб | Частота в популяції | Середнє значення ознаки | Дисперсія |
KK | 70 | 0,07 | 150 | 23,4 |
KL | 300 | 0,30 | 160 | 52,1 |
LL | 400 | 0,40 | 170 | 139,7 |
KM | 200 | 0,20 | 180 | 132,6 |
LM | 20 | 0,02 | 190 | 103,2 |
MM | 10 | 0,01 | 200 | 6,7 |
Загальна дисперсія ……………… . 162,9
Середня середовищна дисперсія … 71,7
Генетична дисперсія …………..….. 91,2
Успадковуваність ………………… 0,56
З таблиці ми бачимо, що дисперсії генотипів значно розрізняються внаслідок їх різної чутливості до умов середовища. Пам’ятаймо, що дисперсія усередині кожного генотипу визначається цілком впливами середовища. Загальна ж мінливість у популяції характеризується величиною загальнопопуляційної дисперсії, яка складається з двох частин. Одна з них – це відмінності між генотипами за їх середніми значеннями ознаки, а друга – відмінності в межах кожного з генотипів, які визначаються тільки середовищем. У нашій популяції генетична складова дисперсії виникає за участю тільки шести генотипів. Звичайно, реальні популяції налічують безліч різноманітних генотипів.
У розглянутій нами гіпотетичній популяції загальна дисперсія виявилася рівною 162,9, а середня середовищна дисперсія – 71,7. Різниця між загальнопопуляційною та середньою середовищною дисперсіями складе генетичну дисперсію популяції – 91,2. Величина генетичної дисперсії означає, що генетична складова обумовлює трохи більше половини всіх індивідуальних відмінностей в популяції.
2.2.5. Показник успадковуваності
Для кращої наочності зобразимо генетичну дисперсію популяції, яку ми аналізували в попередньому розділі (табл. 5), у вигляді долі одиниці, для чого поділимо генетичну дисперсію на загальну: 91,2 : 162,9 = 0,56. Цей показник дістав назву коефіцієнта успадковуваності, який позначається символом h2. Значок квадрата тут вказує на те, що коефіцієнт успадковуваності є складовою дисперсії (σ2). До того ж символом h позначається ступінь домінування.
Таким чином, у розглянутій нами популяції частка генетичної складової у фенотипній дисперсії складає 56%. Це означає, що мінливість даної ознаки в популяції на 56% складається за рахунок мінливості генотипів і на 44% – за рахунок мінливості умов середовища.
Успадковуваність є одним із основних показників, якими оперує генетика кількісних ознак людини, а особливо генетика поведінки. Звичайно цей показник не потрібно сприймати як свідчення чогось неминучого, що неможливо змінити під впливом умов середовища.
По-перше, успадковуваність не є властивістю власне ознаки, а залежить від складу генотипів тієї популяції, на якій проведено дослідження. В іншій популяції з іншим складом генотипів успадковуваність тієї ж ознаки може бути зовсім іншою. Вона може змінитися і тоді, коли одна й та ж ознака буде вивчатися на тій же території, але через деякий час. Адже з часом може змінитися склад генотипів даної популяції внаслідок міграції, вимирання певних генотипів тощо. В обох випадках зміни зазнають як генетична, так і середовищна дисперсії, а отже і коефіцієнт успадковуваності.
По-друге, показник успадковуваності досить чутливий також до змін умов середовища. Повернімося до мал. 17. Зверніть увагу, що в умовах середовища В генетична дисперсія практично відсутня, хоча обидва генотипи наявні. У цьому випадку коефіцієнт успадковуваності близький до нуля. За інших умов середовища (зони А та С) сукупність тих же генотипів показує чітку генетичну дисперсію. Коефіцієнт успадковування у цих випадках буде значимим.
Таким чином, високий показник успадковуваності певної ознаки зовсім не означає, що її не можна змінити під впливом умов середовища. Коефіцієнт успадковуваності свідчить лише про частку генетичної мінливості, яка характерна для даної популяції в даний час та за існуючих умов середовища. Необхідно чітко усвідомити, що він характеризує популяцію, а не конкретного індивіда чи його фенотип. Сучасна генетика не спроможна визначити співвідношення генетичних та середовищних факторів у формуванні фенотипу конкретної особини – людини, тварини і взагалі будь-якого живого організму.
2.2.6. Популяційна мінливість як результат взаємодії генотипу та середовища
Для розуміння природи популяцій важливо пам’ятати, що два основні джерела відмінностей між організмами (генотип і середовище) тісно пов’язано один з одним і перебувають у постійній взаємодії. Відмінності між носіями різних генотипів можуть не виявитися, якщо будуть відсутні ті чинники середовища, які дають змогу генотипним відмінностям стати видимими, фенотипними. Так, люди з винятковими музичними здібностями не зможуть розвинути їх, якщо вони не матимуть нагоди для спеціальних занять. Те ж саме справедливе для будь-якої сфери людської діяльності: тільки в сприятливих умовах середовища люди з особливими природними, спадковими задатками будуть значно відрізнятися від решти особин популяції. Якщо умови середовища не сприятимуть розвитку здібностей, ми навряд чи виявимо великі різницю між фенотипами людей, що мають різні генотипи.
Отже, відмінності між людьми за генотипами можуть виявлятися або не виявлятися залежно від умов середовища: в одному середовищі ці відмінності будуть чітко виражені, тоді як в іншому не з’явиться навіть підозри, що вони існують. Іншими словами, при одному й тому ж генотипному складі популяції можуть існувати умови середовища, які здатні міняти дисперсію фенотипів. Якщо умови середовища ведуть до збільшення фенотипових відмінностей між носіями різних генотипів, результатом буде збільшення фенотипної дисперсії в популяції. Якщо ж середовище нівелює відмінності між індивідами, фенотипна дисперсія зменшуватиметься.
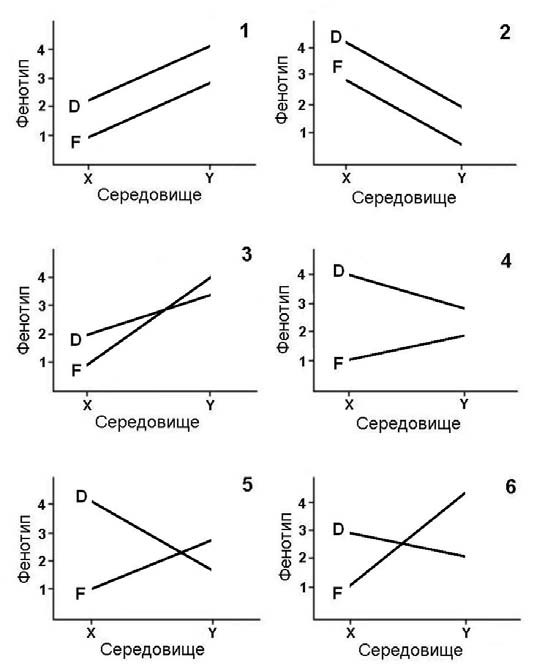
Мал. 18. Приклади генотип-середовищної взаємодії
Це явище дістало назву генотип-середовищної взаємодії. Оскільки така взаємодія впливає на мінливість ознаки в популяції, це означає, що вона здатна формувати певну частку фенотипної дисперсії.
Розберемося з цим на прикладі гіпотетичної популяція, яка складається з двох генотипів – D та F (мал. 18). Вимірювання якоїсь кількісної ознаки цих генотипів здійснюється за різних умов середовища (X та Y). Результати вимірювань залежно від ступеня прояву ознаки позначають цифрами від 1 до 4. Кількість усіх можливих комбінацій чинників D, F, X та Y дорівнює 24 (1·2·3·4 = 24). Але, щоб проілюструвати явище генотип-середовищної взаємодії, достатньо кількох комбінацій. Зрозуміло, що в кожній комбінації аналізується своя ознака та відповідні їй фактори умов середовища.
На перших двох графіках в різних умовах середовища генотип D має вищі значення ознаки, ніж генотип F. Паралельні лінії генотипів означають, що їх фенотипні дисперсії за різних умов середовища залишаються однаковими. Виходить, що генотип-середовищна взаємодія, як складова дисперсії, у даних випадках відсутня.
Перший та другий графіки подібні, хоча розрізняються напрямком зміни фенотипів при переході з середовища X в середовище Y. Для першого варіанту прикладом може бути така ситуація: учні молодших класів, що навчалися у молодого недосвідченого вчителя, потрапляють до сильного педагога, і при цьому всі покращують свої показники, але зберігають ті ж відносні рангові місця. Зворотна ситуація може бути прикладом для другого графіка.
Третій графік відрізняється від перших двох тим, що лінії тут перетинаються. В середовищі X генотип D випереджає генотип F, а в середовищі Y – навпаки. Обидва генотипи при переході з середовища X в середовище Y показують приріст фенотипних значень ознаки, але в різній мірі. Генотип F виявляється чутливішим до умов середовища, бо показує приріст ознаки на 3 одиниці, тоді як генотип D – всього на 1. У цьому випадку генотипсередовищна взаємодія буде відмінною від нуля. Прикладом такої взаємодії може бути така ситуація, коли товариська (D) і нетовариська (F) дитина під час хвороби перебувають в лікарні (середовище X) або удома (середовище Y). Звичайно діти почувають себе краще в домашніх умовах, але для товариської дитини ізоляція від однолітків обтяжує, і їй буде комфортніше в лікарні.
На четвертому графіку генотип D в обох середовищах переважає генотип F, але вони по-різному реагують на зміну умов середовища. При переході з середовища X в середовище Y генотип D зменшує значення ознаки, в той час як генотип F – збільшує. У цьому випадку теж наявна взаємодія генотипу та середовища як частка загальної (фенотипної) дисперсії. Ілюструвати четвертий графік можна ситуацією, коли звичайні (D) та розумово відсталі (F) діти навчаються в звичайній (середовище X) і спеціалізованій (середовище Y) школах. Зрозуміло, що в звичайній школі розумово відсталі діти будуть вчитися значно гірше за звичайних дітей, тоді як в спеціалізованій школі з програмою, розрахованою на розумово відсталих дітей, останні дадуть приріст успішності, а нормальні діти відставатимуть від своїх однолітків.
На п’ятому графіку знову, як і на третьому, спостерігається перетин ліній. При переході з середовища X в середовище Y генотип D демонструє зменшення прояву ознаки, тоді як генотип F – збільшення. До того ж, в середовищі X фенотипні відмінності між генотипами значніші, ніж в середовищі Y. І в цьому випадку генотип-середовищна взаємодія впливає на дисперсію фенотипів. Прикладом розглянутої ситуації можуть бути люди з високим (D) та низьким (F) рівнями активації. В умовах тиші та ізоляції (X) більш ефективно працюватимуть перші, тоді як в умовах шуму (Y) – другі.
Лінії D та F перетинаються і на шостому графіку. Але тут генотипи розрізняються за чутливістю до умов середовища – генотип F реагує на їх зміну значно сильніше, ніж генотип D. Генотип-середовищна взаємодія при цьому суттєва. Шостий графік можна ілюструвати ситуацією, коли одні діти мають середній рівень розвитку з підвищеними просторовими здібностями (генотип D), а інші – яскраві мовні здібності, але слабкі просторові (генотип F). Діти з такими генотипами будуть показувати різну успішність на заняттях з геометрії (X) та літератури (Y).
Розглянуті приклади свідчать, що мінливість у популяції формується не лише за рахунок простого додавання ефектів генотипу та середовища, а й під впливом генотип-середовищної взаємодії.